


INTRODUCTION
Hantavirus and related viruses are the causative agents of Hemorrhagic Fever with Renal Syndrome and Hantavirus Pulmonary Syndrome (HFRS/HPS)1ŌĆō3). Despite extensive studies on etiologic virus and epidemiology, there is no consensus on the pathogenesis of HFRS and HPS. In HFRS/HPS, the major manifestations depend on the vascular change. The change might be the result of the infection of endothelial cells with Hantaan virus, but it is not known to what extent changes in endothelial function are from virus infection or from immune response to virus infection4, 5).
Clinically, there is a varying degree of disseminated intravascular coagulation (DIC) which was evident in the early phase of the illness. It is believed, also, that DIC would be the consequence, at least in part, of functional changes of endothelium resulting in clinical syndrome6). We had reported the possible role of the endotheial cells in the development of HFRS7ŌĆō9).
In inflammatory responses, the expression of various adhesion molecules on the endothelial cells is important. After the initial tethering on the endothelial surface, ICAM-1 expression is believed to be critical in the recruitment and trafficking of mononuclear cells into inflamed lesion through it10, 11). The pathology of HFRS showed a variable degree of inflammatory cell infiltration in various tissues12, 13)
We believed that the vascular endothelial cells would express various kinds of adhesion molecules when they got infected with Hantaan virus and these molecules would play a key role in the establishment of inflammatory lesions in HFRS.
This study investigated the role of adhesion molecule in pathogenesis of Hantaan virus related disease. The expression of ICAM-1 antigen on the cell membrane of human umbilical vein endothelial cells (HUVECs) was assessed by immunohistochemistry, and ICAM-1 mRNA in the endothelial cells was assessed by in situ hybridization after Hantaan virus infection.
METHODS
The materials used in this study were come from the following companies. Gelatin, DEPC, PBS, 20 ├Ś SSC, formamide, paraformaldehyde and HBSS from Sigma (St. Louis, MO, (I.S.A.); FBS and EGM (Endothelial cell growth medium) with 2 % FBS and supplements from Clonetics (San Diego, Ca, U.S.A.); TNF-╬▒ from Biosource (Camarillo, CA, U.S.A.); ICAM-1 monoclonal antibody, In Situ Hybridization Work station and BCIP substrate kit from R&D (Minneapolis, MN, U.S.A.); Anti-mouse alkaline phosphatase immunohistochemistry kit and Fast Red substrate kit from Dako (CA, U.S.A.); Crystal Mount from Biomeda (Foster City, CA, U.S.A.). Plasticwares including Thermanox┬« and glasswares including Superfrost┬« were purchased from Fisher Scientific (Atlanta, GA, U.S.A.).
HUVECs were used in this study. HUVECs had been harvested from a single donor, confirmed by their characteristic expression of vWF Ag and stored after initial propagation in liquid nitrogen until use. The cells were propagated again on the 0.2 % gelatin coated surface with EGM with 15 % FBS. Growth media was fed every 3 days until they reached confluence. For the immunohistochemistry, the cells were transferred to the 0.2 % gelatin coated Thermanox® in the 24-well plates. For the in situ hybridization, cells were propagated on the 0.2 % gelatin coated 6-well plate.
Prototype of Hantaan virus from (Department of Microbiology, College of Medicine, Chung-Ang University, Seoul, Korea) was infected on the HUVECs. Virus was propagated on the Vero E-6 cells for 13 days. After clearing of culture supernatant by centrifugation, virus solution was stored in liquid nitrogen until use. The infectivity of the virus solution was assessed by classic plaque assay.
Virus was adsorbed with the dose of 0.2 ŌĆō 0.5 PFU/HUVECs for 60 min. after washing twice with HBSS. After removal of remained virus solution, cells were fed with maintenance media (growth media with 5 % FBS) and maintained for 3 hours, 6 hours, 12 hours, 1 day, 2 day and 3 days, respectively.
The virus solution irradiated for 5 ├Ś 105 rad was treated in same way to prepare the negative control, and TNF-╬▒ was added into media in the concentration of 5 ng/mL for the positive control.
The cells were fixed at once with cold 4 % paraformaldehyde in PBS for 20 min. Before the fixation, the cells on the Thermanox┬« were washed twice with HBSS and fixed with an ample amount of fixative. The Thermanox┬« were dehydrated in graded alcohol and stored in ŌłÆ70┬░C until use. For the immunohistochemistry, cells were rehydrated in graded alcohol and kept in PBS for 10 min.
For immunohistochemistry, anti-ICAM-1 monoclonal antibody was reacted in 1:3,000 dilution with 5 % skim milk in PBS for 16 hours at 4ŌĆ▓C. After a wash with PBS ŌłÆ 0.2 % Tween 20 twice, 300 uL of link antibody was applied for 30 min. at room temperature. After a wash with PBS-Tween 20 twice again, 300 uL of alkaline phosphatase conjugate was applied again and reacted for 30 min. at 37┬░C. For the coloring reaction, Fast Red substrate was applied for 5 to 15 min. along the intensity of color reaction. Each Thermanox┬« was fixed on the glass slide and mounted with Crystal Mount after the counter-staining with MeyerŌĆÖs hematoxylin.
For in situ hybridization, the cells were trypsinized from the 6-well plates and washed with RNAse free PBS twice. Cells were fixed for 20 min. with cold 4 % paraformaldehyde and cells were washed with RNAse free 3 ├Ś PBS once and 1 ├Ś PBS twice. Cells were suspended in appropriate volume of RNAse free PBS and smeared on the Superfrost┬« slides. After drying for an hour, slides were treated with 2 ├Ś SSC and DEPC water briefly. The cells were reacted with 300 uL of prehybridization/hybridization solution for 60 min. at 37┬░C. Remaining solution was removed and cells were reacted with 300 uL of digoxigenin labeled ICAM-1 probe cocktail in the concentration of 200 ng/mL for 16 hours at 42ŌĆ▓C. After serial wash with 4 ├Ś SSC/30 % formamide, 4 ├Ś SSC/30 % formamide and 0.2 ├Ś SSC/30 % formamide twice, respectively, slides were washed with modified TBS (50 mM Tris, pH 7.6, 150 mM NaCl, 2 mM MgCl2, 0.1 % BSA and 0.1 % Triton). Slides were reacted with sheep anti-digoxigenin/alkaline phosphatase conjugate for 30 min. at room temperature in the dilution of 1:600. After a wash with modified TBS twice, 300 uL of revealing agent (substrate 5-bromo-4-chloro-3-indolylphosphate (BCIP) was overlayed and reacted for 8 hours at 37┬░C. Slides were mounted with Crystal Mount after a brief rinse with distilled water and countersigned with 0.1 % Malachite Green.
To improve the preparation, the above procedure was repeated on the cells grown on the Thermanox® 12 hours after Hantaan virus inoculation, 6 hours after TNF-stimulation in the concentration of 200 ng/mL and 12 hours after radiation sterilized Hantaan virus inoculation, respectively.
RESULTS
1) Determination of Hantaan Virus Infectivity
By classic plaque assay, the infectivity of our virus solution was calculated as the following equation and it was 2.6 ├Ś 104 PFU/mL. We could count plaques clearly when we had overlaid Neutral Red-Noble Agar on day 13 and waited for 2 days. (Fig. 1)
2) Immunohistochemistry against Hantaan antigen
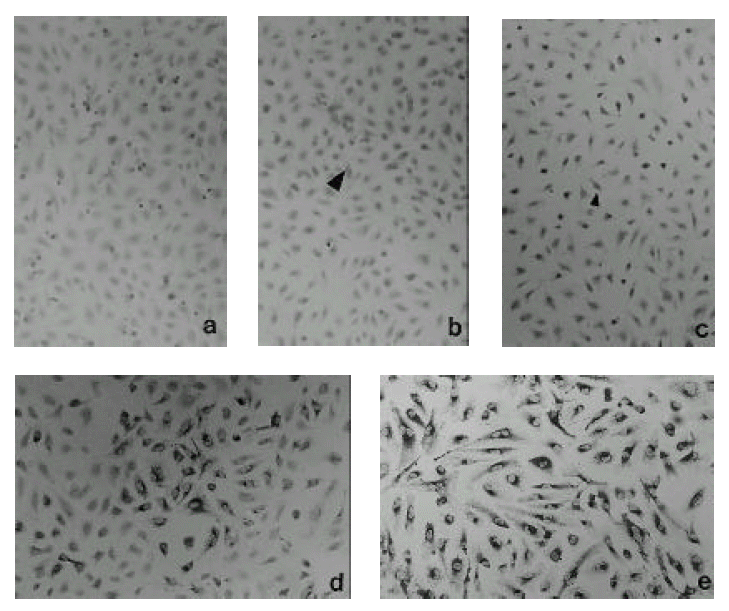
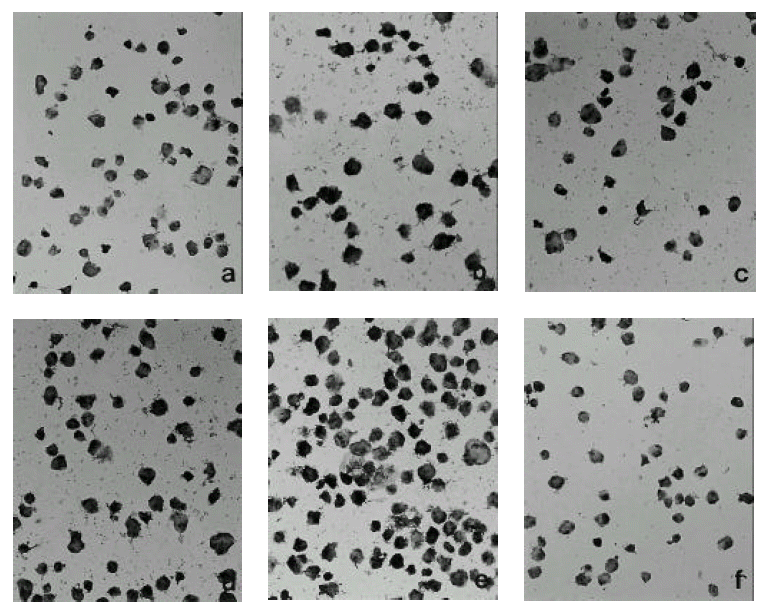
We could not find any positive cells when we had inoculated radiation sterilized virus solution. (Fig. 2-a) With Hantaan virus inoculation, we could find a small number of scattered positive cells throughout the cell layer after 12 hours. (Fig. 2-b) After 24 hours, the positive islands against Hantaan antigen increased in number and started to distribute all over the monolayer (Fig. 2-c). Three days after inoculation, the positive cells increased markedly in their number and distribution (Fig. 2-d). On day 7, almost all cells were demonstrated to have Hantaan antigen by immunohistochemistry. (Fig. 2-e)
3) Immunohistochemistry against ICAM-1
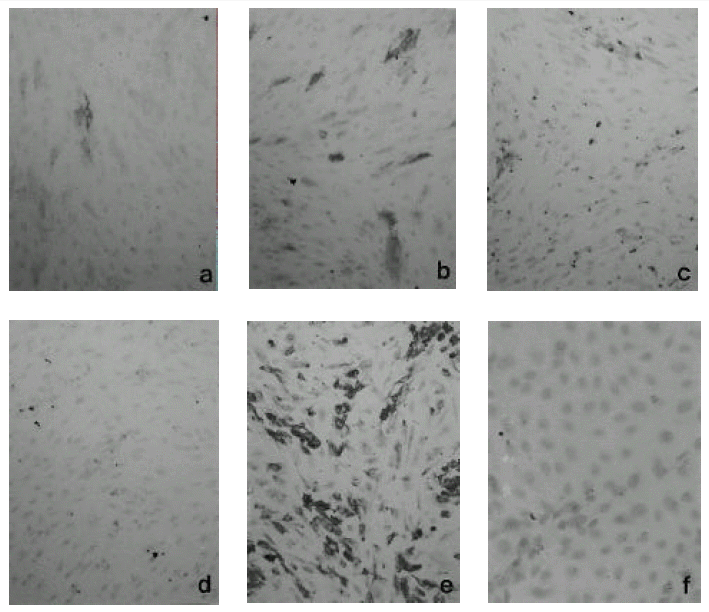
Hantaan virus inoculation expressed ICAM-1 antigen after 6 hours and ICAM-1 expression increased with time. (Fig. 3-a) ICAM-1 antigen expressed high between 12 hours and 24 hours of postinoculation. The percentage of positive cells is between 5 % and 10 %. (Fig. 3-b) After day 1, the expression decreased abruptly and was barely seen in the day 4 specimen. (Fig. 3-c & d)
4) In Situ hybridization against the mRNA of ICAM-1
When Hantaan virus was inoculated, we could observe positive reaction after 4 hours that increased in intensity and extent along the course. The positive reaction was evident between day 1 and day 2 and tended to decrease (Fig. 4-a, b, c & d).
We used TNF-╬▒ stimulated HUVEC for 8 hours in the concentration of 200 ng/mL as positive control (Fig. 4-e). As negative control, we employed radiation sterilized Hantaan virus and did hybridization 12 hours later. (Fig. 4-f)
In the cells grown on the Thermanox®, the expression of mRNA of ICAM-1 was evident in comparison to that of radiation sterilized virus infected cells and the number of positive cells was comparable to that of TNF-α stimulated cells. (Fig. 5-a, b & c)
DISCUSSION
Hantavirus and related viruses are the causative agents of HFRS and HPS. Despite extensive clinical-epidemiological and virologic studies, there is no consensus on the pathogenesis of HFRS and HPS.
The agent designated Hantaan virus is a single stranded RNA virus belonging to the family Bunyaviridae, genus Hantavirus. The genus consists of at least five species; Hantaan virus (Korean Hemorrhagic Fever), Seoul virus, Puumala virus (Nephropathia Epidemica), Porogia virus (Balkan HFRS) and Prospect Hill virus14, 15)
In the PeopleŌĆÖs Republic of China, over 100,000 cases of HFRS are reported annually and the incidence is increasing. Hundreds of cases of nephropathia epidemica have occurred annually in Finland and other Scandinavian countries since the 1930ŌĆÖs. Antibody studies indicate worldwide distribution. Antibodies in human sera have been found in South and North America (including Hawaii and Alaska), Southeast Asia and Africa. Furthermore, laboratory acquired cases have been reported in Russia, Korea, Japan and Europe, with rats implicated in Korea, Japan and the United Kingdom15, 16). The mortality rates are between 3 to 32 percent in the former Soviet Union and between 7 and 15 percent in China. During the outbreak in the New Mexico-Arizona-Colorado region, two-thirds of the cases were fatal. Between April 1951 and December 1976, the overall case fatality ratio in Korea was 6.6 percent15).
Although it is not clear how the clinical syndrome, including fever, shock, bleeding tendency, renal failure and hypopituitarism happens in HFRS, it is believed that the basic functional disturbances depend on the vascular changes.
The vascular changes in HFRS may be a result of the infection of endothelial cells with Hantaan virus, but it is not known to what extent changes in endothelial function are from virus infection or from immune response to virus infection. There is no doubt that there is variable degree of DIC in the early phase of HFRS, Clinically, DIC was evident in all patients who visited the hospital 5 days from the onset of the illness17, 18). DIC is believed to be a consequence, at least in part, of functional changes of endothelium resulting in kinin activation and clinical syndrome6, 9). The most plausible initiating mechanism of DIC is the exposure of pro-coagulant activity from the endothelial cells by the infection of the virus or from the endothelium damaged with immune complex or platelet-anti-platelet antibody complex.
We had reported the possible role of the endothelial cells in the development of HFRS. We were able to localize viral antigens in the endothelial cells and demonstrated the changes of anti-coagulant characteristics of endothelial cell membrane after Hantaan virus infection7ŌĆō9)
Pathology also showed edema and destruction and other evidence of injury on the endothelial cells of small vessels in HFRS12). Hantaan antigen also had been found in the endothelial cells of brain, lung, heart, liver and kidney of autopsy cases9, 13).
The expression of various adhesion molecules on the endothelial cells are important in the initiation of inflammatory reaction and its propagation into adjacent tissues. For the establishment of inflammation in specific tissue, there should be tropism between circulating inflammatory cells and vessels in those tissues. It should help inflammatory cells to be localized in a specific area. ICAM-1 expression is critical in the recruitment and trafficking of mononuclear cells into inflamed lesion through it11). The pathology of HFRS showed a variable degree of inflammatory cell infiltration in various tissues. The kidney is infiltrated heavily with mononuclear cells along with the destruction of tubular epithelial cells and medullary interstitial hemorrhage and varying degrees of vascular changes12, 13).
We believed that the vascular endothelial cells would express various kinds of adhesion molecules when they got infected with Hantaan virus, and these molecules would play a key role in the establishment of inflammatory lesions in HFRS.
Three members of the immunoglobulin gene family are expressed on endothelial cells and are involved in leukocyte emigration and activation, ICAM-1, ICAM-2, and VCAM-1, with a fourth member, ICAM-3, expressed on leukocytes. ICAM-1, ICAM-2 and ICAM-3 account for all known binding of LFA-1 to cells. The differences in tissue distribution of the three LFA-1 ligands suggest that it is unlikely their functions completely overlap. Binding affinity differs among the different ICAM-species, with ICAM-2 and ICAM-2 binding to LFA-1 expressing cells more weakly than ICAM-1. Expression of ICAM-1 was found to be markedly elevated in response to interleukin 1 (IL-1), tumor necrosis factor (TNF), interferon-╬│ (IFN-r) and bacterial lipopolysaccharide (LPS) in a wide variety of cells, including endothelial cells. ICAM-1 and/or ICAM-2 is thought to be responsible for the basal binding of lymphocytes to unstimulated HUVECs, indicated by the ability of MoAb against CD18 to inhibit this binding almost completely19).
In this study, the changes of ICAM-1 were investigated on the Hantaan virus infected HUVECs by immunohistochemistry and in situ hybridization.
ICAM-1 expression was evident in the early hours of infection on HUVECs. The number of ICAM-1 positive cells increased with time during the 12 or 24 hours after infection. Although there was some variation in the proportion of positive cells, 5 to 10 % of HUVECs were positive after 12-24 hours. The number of positive cells exceeded by far in comparison to gamma irradiated virus. They had been employed as negative control. They distributed in patch and the number of ICAM-1 positive cells increased and reached the peak after 12ŌĆō24 hours of post-inoculation.
As would be expected by the broad tissue distribution of ICAM-1, these protein play a more general role in the immune response than just emigration of leukocytes out of the vasculature. ICAM-1 plays an accessory, antigen independent role in these adhesive interactions. One such accessory role in which ICAM-1 functions is to enhance antigen dependent T cell activation. The presence of ICAM-1 on the surface of the antigen presenting cell results in a shift in antigen dose response curves, such that the T cell becomes activated by 10 to 100-fold lower concentrations of antigen20). ICAM-1 may also provide coactivation signals for bacterial superantigen activation of T lymphocytes21). Other activities which may be involved include T cell dependent B cell activation, cytotoxic T lymphocyte mediated killing of target cells, natural killer cell mediated killing of target cells, neutrophil induced oxidative damage of target cell and T lymphocyte development. VCAM-1 was known to elicit a co-stimulatory signal to resting T cells.
When Hantaan virus infected endothelial cells up-regulated the expression of ICAM-1 and, possibly, other adhesion molecules, they could recruit the circulating mononuclear cells and platelets. Endothelium and attached mononuclear cells and platelets could provide a vast area of phospholipid surface and it could lead into the activation of a coagulation system and DIC. Recruiting mononuclear cells from the circulation would express tissue factor activity along with the infection of the endothelium, and it would contribute to the establishment of DIC. It deserves further study.
This study has a few pitfalls. We did not pursue any dynamic change. In vivo, stimulated endothelium can express adhesion molecules that increase the adhesion of monocytes and results in the production of IL-1, TNF and TF. TNF-╬▒ increases the expression of adhesion molecules and recruits mononuclear cells upon the endothelial cells. The recruited cells are able to express tissue factor and they can amplify the coagulation process, too. They may secure the concentration of cytokine high in the lesion and can influence mononuclear cell adhesion in various ways and induce pro-coagulant activity. We did not study the kinetics of various cytokines and their effects on the HUVECs infected with Hantaan virus. It may establish the local effect and response of HUVECs.
In conclusion, the human umbilical vein cells infected with Hantaan virus express ICAM-1 for circulating inflammatory mononuclear cells, and it may contribute organ specific inflammatory processes, including kidney, heart and CNS in HFRS.


 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print





