


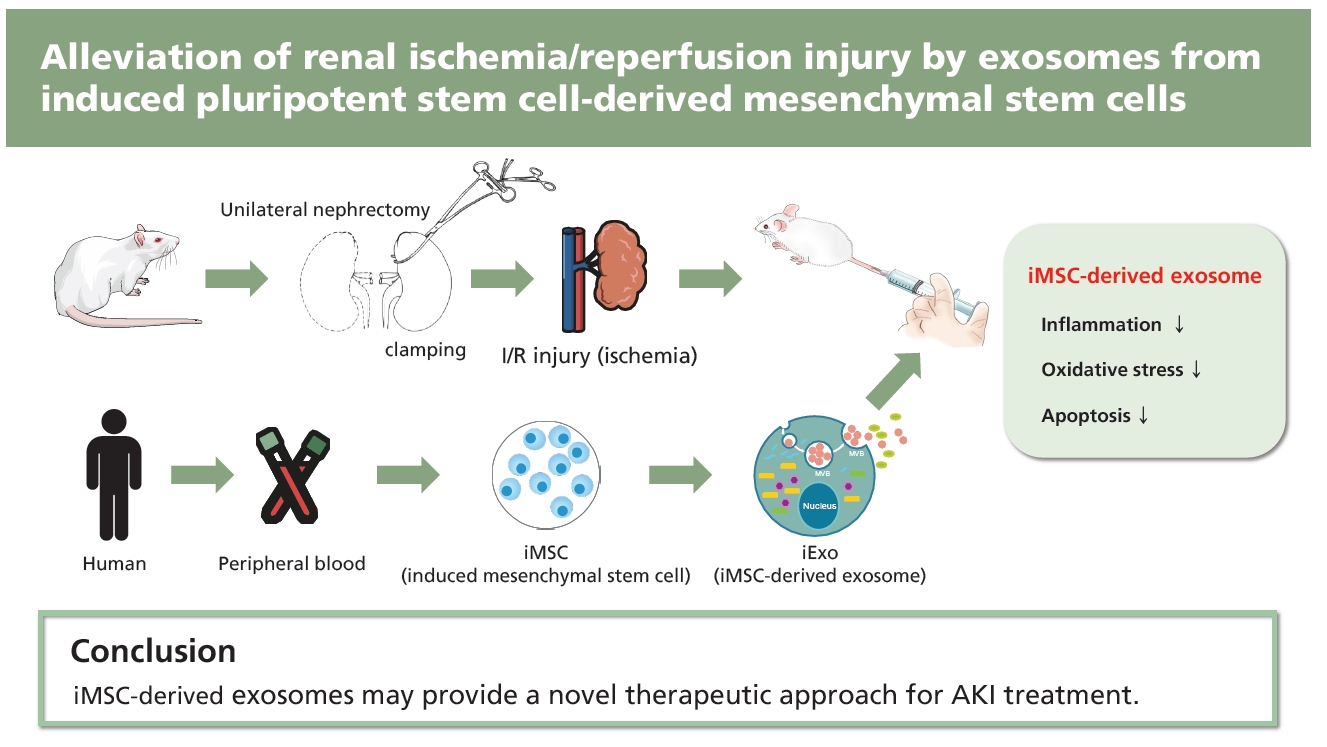
INTRODUCTION
Renal ischemia followed by ischemia reperfusion (I/R) is a leading cause of acute kidney injury (AKI) and acute kidney failure, which are associated with high morbidity and mortality. Previous studies have shown that ischemia is related to epithelial cell necrosis and endothelial cell dysfunction; reperfusion can also result in renal tubular cell apoptosis [1–4]. Acute tubular necrosis induced by I/R injury severely affects kidney graft outcomes in transplanted kidneys [3,5–7]. Therefore, I/R injury is a critical clinical challenge in both native and transplanted kidneys, with limited effective treatments available.
Recent advances in stem cell technology have enabled the generation of patient-specific induced pluripotent stem cells (iPSCs) from adult somatic cells, which can differentiate into expandable progenitor cells and mature cells [8]. iPSCs exhibit similar properties with embryonic stem cells (ESCs) with self-renewal and differentiation properties. However, one distinct advantage over ESCs is that iPSCs are patient-specific and, therefore, theoretically, can overcome the need for immunosuppression in the recipient. iPSCs can generate unlimited amounts of early passage patient-specific mesenchymal stem cells (MSCs) with consistent quality. iPSC-derived MSCs (iMSCs) provide a promising cell source for autologous cell therapies in regenerative medicine because of their more powerful therapeutic function compared to bone marrow-derived mesenchymal stem/stromal cells (BMSCs) [9,10].
Although it has been demonstrated that MSCs exhibit advantages in cell therapy, one potential challenge is their acquisition of genetic and epigenetic alterations. MSCs become immortalized after long-term culture and can spontaneously transform into malignant cells due to enhanced chromosomal instability associated with dysregulation of telomere activity and cell cycle-related genes, resulting in tumorigenesis when injected into multiple organs [11]. Jeong et al. [12] found that even short-term cultured MSCs transplanted into mice can transform into malignant tumors. Therefore, determining how to fully advantageously use MSCs while avoiding the adverse effects, such as tumor formation, is an important step toward their application in the clinical setting. Recently, accumulating evidence has indicated that the therapeutic action of stem cells is mainly exerted through the secretion of molecules such as growth factors, cytokines, chemokines, and extracellular microvesicles into the surrounding environment via a paracrine mechanism [13]. Among these paracrine molecules, exosomes demonstrate unique functions in disease diagnosis and therapy [14,15]. Exosomes are small membrane vesicles (40 to 100 nm diameter) of endosomal origin formed in multivesicular bodies (MVBs) by invagination of the endosomal membrane and then released into the extracellular space when MVBs fuse with the plasma membrane [16].
Recent studies have indicated that exosomes derived from BMSCs can promote angiogenesis in ischemic tissue and attenuate tissue damage after ischemic injury [17–19]. Further, it has been confirmed that exosomes derived from MSCs are immune-tolerant, an important property for clinical applications [20]. Therefore, we hypothesized that exosomes derived from iMSCs (iExo) may similarly reduce tissue injury after I/R. In the present study, we investigated the therapeutic effect of iExo in a mouse renal I/R model, demonstrating important implications for new therapeutic approaches to AKI.
METHODS
Generation of human iPSCs from peripheral blood mononuclear cells
Peripheral blood samples were obtained from healthy human donors after obtaining informed consent, with approval of the Institutional Review Board of Seoul St. Mary’s Hospital (KIRB-00613_6-001). Peripheral blood mononuclear cells (PBMCs) were isolated via centrifugation using Ficoll-Paque PLUS media (GE Healthcare Bio-Sciences AB, Uppsala, Sweden). Isolated PBMCs were cultured in StemSpan ACF (Stem Cell Technologies, Vancouver, BC, Canada) supplemented with StemSpan CC110 for 4 days. Mononuclear cells were then transferred to a 24-well vitronectin-coated plate (BD BioCoat, BD Biosciences, Franklin Lakes, NJ, USA), and Sendai virus (SeV) (CytoTune-iPSC 2.0 Reprogramming kit, Thermo Fisher Scientific, Waltham, MA, USA) was added to give a multiplicity of infection of three (MOI:3). The medium was changed daily until iPSC colonies formed. After manual picking, iPSC lines were maintained on vitronectin-coated plates (Thermo Fisher Scientific) in a TeSR-E8 medium (Stem Cell Technologies). Emerging iPSC colonies were picked individually and expanded for characterization on day 12 after transduction. Cells were cultured in a 37°C incubator with 5% CO2 from days 3 to 21 after transduction.
Generation of MSCs from iPSCs
To reprogram iPSCs into iMSCs, iPSCs were dissociated with the aid of Versene thylene-diamine-tetraacetic acid (Thermo Fisher Scientific) and seeded into nonadherent 6-well conical polymerase chain reaction plates (10,000 cells per well) with Iscove’s Modified Dulbecco’s Medium (MDM basal media, 17% KnockOut Serum Replacement, 1% minimal essential medium nonessential amino acids, 110 mM 2-mercaptoethanol, and 1% PSA antifungal-antibacterial solution; Thermo Fisher Scientific). On day 2, embryoid bodies (EBs) were transferred to nonadherent 2.4 mg/cm2 poly-HEMA-coated flasks and cultured for 3 more days. On day 5, EBs were transferred to 1% gelatin-treated flasks for 3 more days. On day 8, some EBs were attached to the surface and cells grew outward from the EBs. Non-attached EBs were transferred to another 1% gelatin-coated flask. Cells were fed with medium supplemented with 10 ng/mL TGF-β1 (R&D Systems, Minneapolis, MN, USA) on days 8 to 10, after which the medium was switched to standard Dulbecco’s Modified Eagle Medium culture medium containing 10% fetal bovine serum, 2 mM L-glutamine, and 100 U/mL penicillin/streptomycin (Thermo Fisher Scientific). The medium was changed twice weekly, and cells were split upon confluence in a 1:3 ratio. The same iMSC derivation protocol was repeated using three different iPSC control lines.
Isolation and purification of exosomes from iMSC
After iPSC-MSCs reached approximately 80% confluency, the culture medium was replaced with MesenGro hMSC medium (StemRD Inc., Burlingame, CA, USA), and cells were cultured for an additional 48 hours. Conditioned medium (CM) was collected, and exosomes were isolated as previously described [21]. Briefly, the CM was centrifuged sequentially at 300 g for 10 minutes and then at 2,000 g for 10 minutes. Next, cellular debris in CM supernatants was removed via 0.22-μm filtration, and supernatants were ultracentrifuged at 100,000 g for 2 hours. Pelleted exosomes were resuspended in phosphate-buffered saline and then centrifuged at 4,000 g until the volume in the upper compartment was reduced to approximately 200 μL. Total protein concentration in exosomes was quantified using the Micro Bicinchoninic Acid Protein Assay Kit (Thermo Fisher Scientific), according to the manufacturer’s recommended protocol. All procedures were performed at 4°C. Exosome morphologies were observed using a JEOL 1200EX transmission electron microscope (TEM; JEOL Ltd., Tokyo, Japan). Primary antibodies against tumor susceptibility gene 101 (TSG101) (Cat#SC-136111, 1:200; Santa Cruz Biotechnology, Dallas, TX, USA), CD9 (Cat#SC-13118, 1:200; Santa Cruz Biotechnology), CD63 (Cat#SC-365604, 1:200; Santa Cruz Biotechnology), and secondary antibody were added. Exosomal marker proteins were used to analyze the incorporation of each protein into exosomes in immunoblots.
Flow cytometric analysis
iMSC surface antigens were analyzed using a FACSCalibur flow cytometer (BD Biosciences) employing FlowJo software version 10 (Tree Star, Ashland, OR, USA). The following conjugated monoclonal antibodies (BD Biosciences) were used at concentrations recommended by the manufacturer: CD34-phycoerythrin (PE) (Cat#12-0349-42), CD31-fluorescein isothiocyanate (FITC) (Cat#11-0319-42), CD19-allophycocyanin (APC) (Cat#152409), CD11-FITC (Cat#11-0112-41), human leukocyte antigen–DR isotype (HLA-DR)-FITC (Cat#11-9956-41), CD44-FITC (Cat#11-0441-81), CD73-FITC (Cat#11-0739-41), CD105-FITC (Cat#12-1051-81), and CD90-FITC (Cat#11-0909-41). We also prepared appropriate isotype staining controls.
I/R injury modeling and iMSC or iExo injection
Animal study protocols were approved by the Institutional Animal Care and Use Committee at the Catholic University of Korea (CUMC-2017-0245). AKI was induced by kidney I/R in 8-week-old male BALB/c mice. I/R injury was induced by unilateral renal pedicle clamping plus contralateral nephrectomy, as previously described [22]. After front incisions were made, blood supply to the kidneys was blocked for 25 minutes. Saline, iMSCs (1 × 105 or 2 × 105/mouse), or iExo (10 or 50 or 200 μg/mouse) were injected through the tail vein after reperfusion. Animals were euthanized after 24 hours of reperfusion. Control animals were subjected to sham operations without renal pedicle clamping.
Measurement of serum creatinine
Serum creatinine (sCr) was evaluated by a quantitative enzymatic colorimetric method (0430-120; Stanbio Laboratory, Boerne, TX, USA), according to the manufacturer’s instruction.
Histopathological examination
Mouse kidneys were fixed in 4% paraformaldehyde and embedded in paraffin, and 4-um sections were cut and stained with periodic acid-Schiff stain (PAS, Sigma-Aldrich, St. Louis, MO, USA). Renal tubular damage was scored by a blinded renal pathologist. A grade scale of 0 to 5, as outlined by Lien et al. [23], was used for the histopathological assessment of I/R-induced renal injury. Histological changes due to tubular necrosis were quantified by calculating the percentage of tubules displaying acute tubular necrosis (ATN), loss of brush border, cast formation, and tubule dilatation: 0, no damage; 1, < 25% damage; 2, 25% to 50% damage; 3, 50% to 75% damage; 4, 75% to 90% damage; and 5, > 90% damage. At least 10 fields (original magnification ×100) were reviewed in each slide.
In situ terminal deoxynucleotidyl transferase dUTP nick end labeling
Apoptotic cells in mouse kidney tissue sections were detected using the in situ terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) method and stained with the In Situ Apoptosis Detection Kit (Cat#MER-S7100; MERCK-Millipore, San Diego, CA, USA). TUNEL-positive cells were counted in approximately 20 randomly selected, non-overlapping areas per animal in each group.
Quantitative real-time polymerase chain reaction
Total RNA from mouse kidney tissues was isolated using an RNA-spin Total RNA Extraction Kit (iNtRON Biotechnology Inc., Seongnam, Korea). First-strand cDNA was synthesized using a Reverse Transcription System (Promega, Madison, WI, USA) and subjected to quantitative real-time polymerase chain reaction (qRT-PCR) using SYBR Green Master Mix in the LightCycler 480 system (Roche Life Science, Rotkreuz, Switzerland). Gene expression was normalized to glyceraldehyde 3-phosphate dehydrogenase (GAPDH) expression using the change-in-threshold method and primers with the following sequences: 5′-GGC CAA GGG AGA TGT TAC AAC-3′ and 5′-GCA ACT CTC CTT TGG GTT CTC-3′ for mouse superoxide dismutase 2 (SOD2), 5′-GCG GAT TCC TGA GAG AGT GGT AC-3′ and 5′-GCC TGA CTC TCC AGC GAC TGT GGA G-3′ for mouse catalase, 5′-CAC ATT CCC AAA CAA GAT GC-3′ and 5′-TCT TTT TCC AGC GAG GAG AT-3′ for mouse nuclear factor erythroid-2-related factor (NRF2), 5′-CTT CCT CAG CCA TGG TAC CTC T-3′and 5′-CAA GTC TTC ATC AGC ATC AAA CTG-3′ for mouse nuclear factor κB (NF-κB), 5′-CTT CTC ATT CCT GCT TGT G-3′ and 5′-ACT TGG TGG TTT GCT ACG-3′ for mouse tumor necrosis factor-α (TNFα), 5′-TTC TTC AGC AAC AGC AAG GC-3′ and 5′-CCT TTT CCT CAG CGA CGA CT-3′ for mouse interferon γ (IFNγ), 5′-TTG TGG CTG TGG AGA AG-3′ and 5′-CAT CAG AGG CAA GGA GG-3′ for mouse interleukin 1β (IL-1β), 5′-CCC ATG TTT GTG ATG GGT GT-3′ and 5′-GTG ATG GCA TGG ACT GTG GT-3′ for mouse GAPDH [22,24].
Detection of 8-OHdG in urine
An enzyme-linked immunosorbent assay (ELISA) kit (Cell Biolabs, San Diego, CA, USA) was used to detect 8-OHdG in spot urine, according to the manufacturer’s instructions. Urinary 8-hydroxy-2′-deoxyguanosine (8-OHdG) concentration was calculated as picogram per milligram of creatinine.
Immunohistochemical analysis of 8-OHdG in tissue sections
Immunohistochemistry was carried out as described above [24]. Briefly, 4-μm sections of paraffin-embedded kidney tissue were deparaffinized and then placed in citrate buffer solution (pH 6.0), methanolic H2O2, and 0.5% Triton X-100. Nonspecific binding sites were blocked in 10% normal donkey serum (Jackson ImmunoResearch, West Grove, PA, USA). Sections were incubated overnight at 4°C with anti-8-OHdG antibodies (Cat#MOG-100P, JaICA, Shizuoka, Japan) and then with peroxidase-conjugated secondary antibodies for 2 hours at room temperature. Peroxidase activity was detected using 3,3′-diaminobenzidine (Vector Laboratories, Burlingame, CA, USA) as a chromogen. Quantitative analysis was performed by calculating the 8-OHdG positive area showing the same intensity, using histogram equalization (TDI Scope Eye version 3.6 for Windows, JN OpTIC, Seoul, Korea).
Immunoblot analysis
Whole kidneys were lysed in 10 mM Tris (pH 7.5) containing 1% sodium dodecyl sulfate and 1 mM NaVO4. Equal amounts of protein were subjected to immunoblot analysis using the following primary antibodies from Cell Signaling Technology: phosphorylated-mitogen-activated protein kinase kinase 1/2 (p-MEK1/2; Cat#9121), total-MEK1/2 (Cat#8727), p-EKR1/2 (Cat#9101), and total-ERK1/2 (Cat#9121). Signals were detected using an enhanced chemiluminescence system (ATTO Corp., Tokyo, Japan). Quantification of relative intensities was performed with the sham group set as 100%; densities were normalized to that of total form bands from the same gel using Quantity One version 4.4.0 software (Bio-Rad Laboratories, Hercules, CA, USA).
Statistical analyses
Data were expressed as the mean ± standard error from at least three independent experiments. Multiple comparisons between groups were performed via a one-way analysis of variance with Bonferroni post hoc testing using Prism software version 7.03 for Windows (GraphPad Software, La Jolla, CA, USA). Results with p values < 0.05 were considered significant.
RESULTS
Characterization of iMSCs and iExo
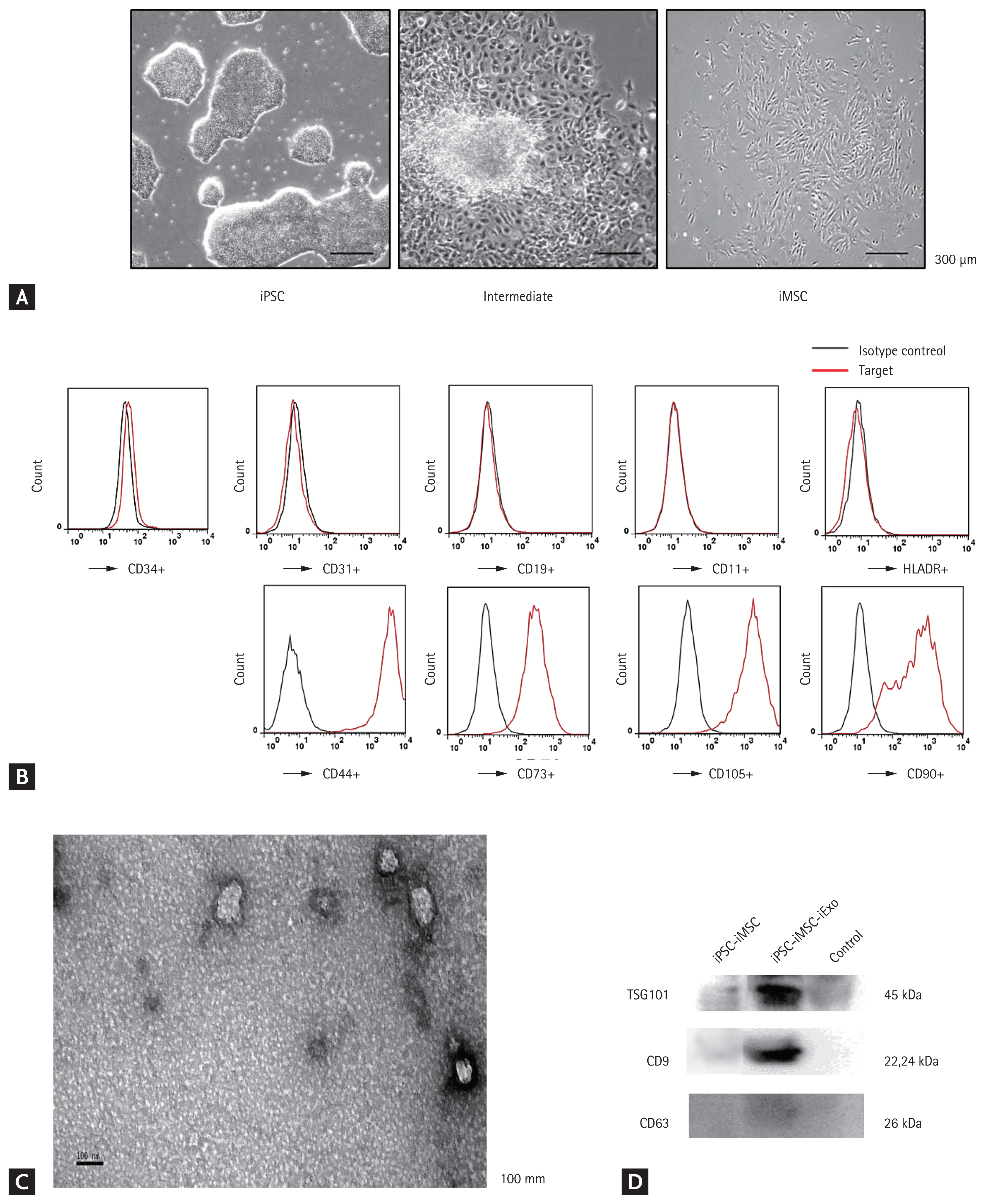
We successfully derived human MSCs from three different iPSC cell lines using an induction protocol. After culturing in MSC medium for a few days, iPSCs showed a tendency to form packed clones with decreased nuclear:cytoplasmic ratios and formed a monolayer with a larger spindle-shaped morphology at the border of the colonies. After culturing on gelatin-coated dishes for 14 days, cells were continually passaged when they reached 90% confluence until homogeneous fibroblastic morphologies were observed (Fig. 1A). A longitudinal study comparing MSC surface marker expression at passage 5 was conducted using flow cytometry to monitor the transition of iPSCs to the MSC-like phenotype (Fig. 1B). After a few passages in culture, iMSCs expressed similar levels of CD44, CD73, CD105, and CD90 as BMSCs. In TEM experiments with iExo, we observed spheroidal microvesicles that were 30 to 100 nm in diameter (Fig. 1C), indicating the presence of exosomes. Western blot analyses indicated that iExo expressed exosomal markers such as TSG101, CD9, and CD63 proteins (Fig. 1D).
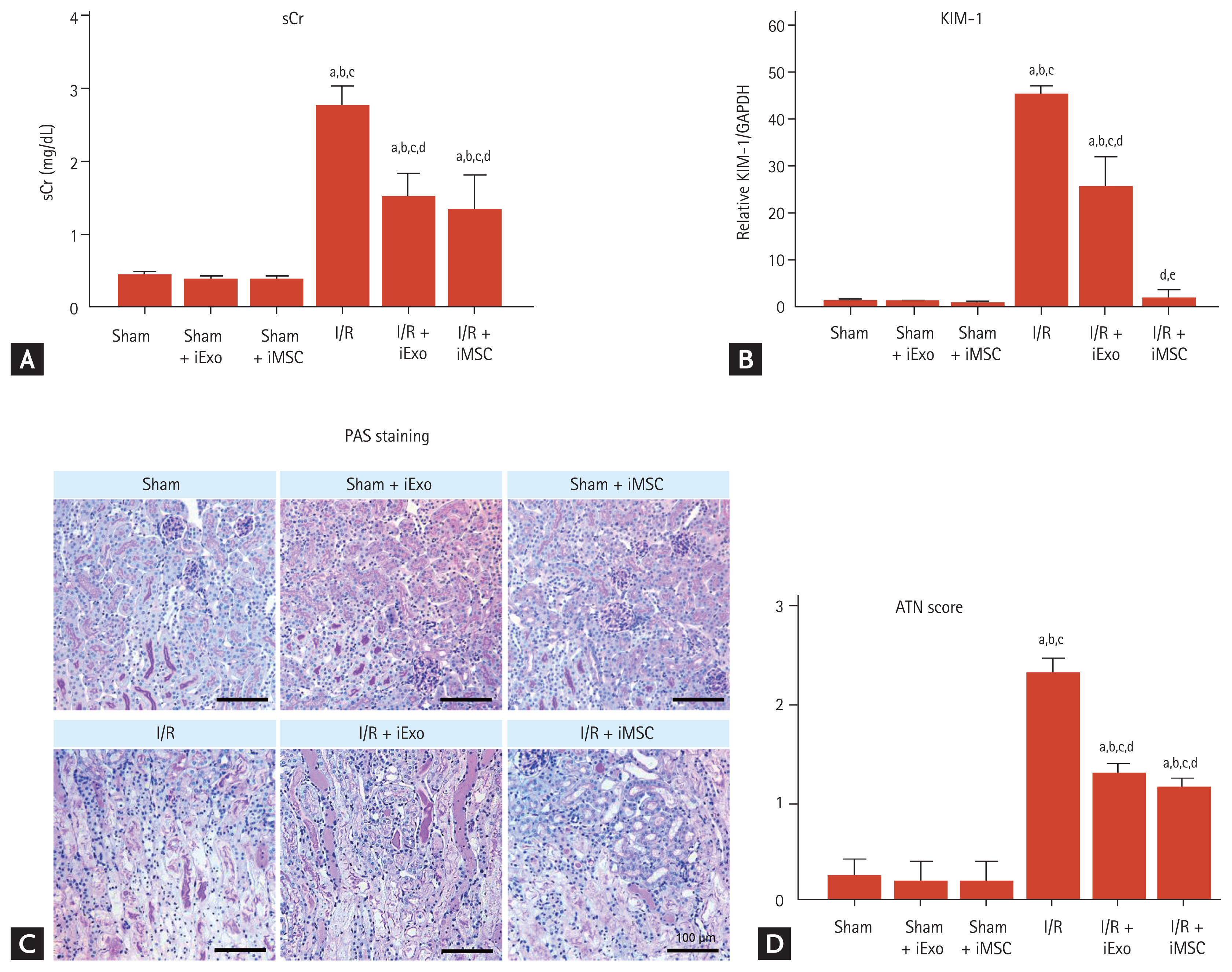
Effects of treatment with iExo or iMSCs on renal function and histology in renal I/R injury mouse model
Mice with I/R injury in the iExo treatment (I/R + iExo) group had significantly lower sCr levels and ATN scores after 24 hours compared to mice in the I/R group, as shown in Fig. 2 (sCr, 1.53 ± 0.31 mg/dL vs. 2.77 ± 0.25 mg/dL; ATN score, 1.3 ± 0.11 vs. 2.3 ± 0.15, p < 0.05 vs. the I/R group). Mice treated iMSCs had significantly reduced sCr levels and ATN scores compared to those in the I/R group but did not differ significantly from those in the I/R + iExo group (sCr, 1.35 ± 0.45 mg/dL; ATN score, 1.16 ± 0.09, p > 0.05 vs. the I/R + iExo group). The sham groups that received iMSC or iExo injections did not differ significantly.
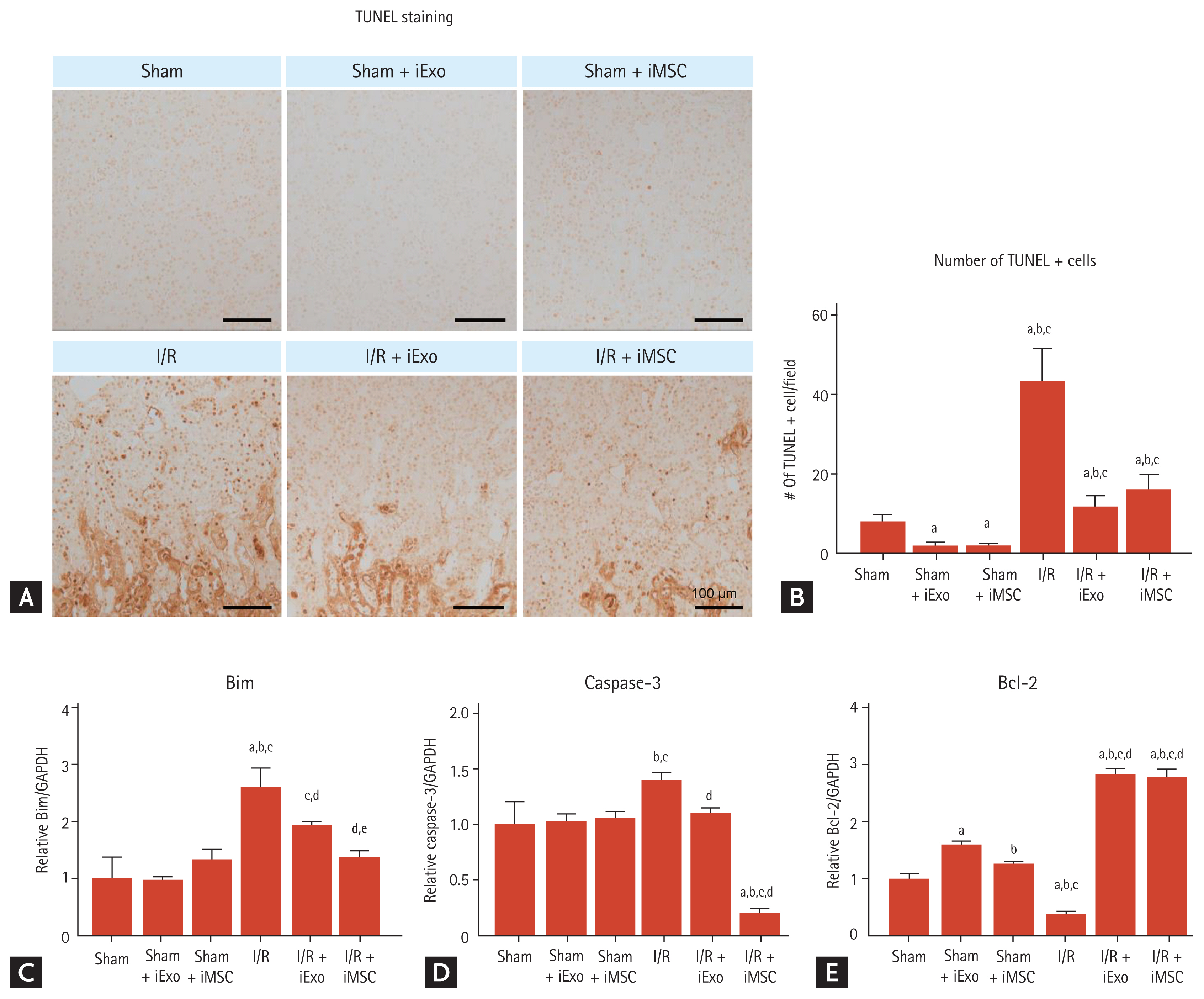
Effects of iExo or iMSC treatment on apoptosis induced by renal I/R injury
Next, we investigated whether iMSC or iExo treatment had any effect on the apoptotic process during I/R injury. The sham groups exhibited few TUNEL-positive cells, but treatment with iExo or iMSCs resulted in significantly fewer TUNEL-positive cells (1.8 ± 0.9 and 1.8 ± 0.5 vs. 8.0 ± 0.2, p < 0.05 vs. the sham group). In the I/R group, numerous cells were strongly positive for TUNEL staining, as shown in Fig. 3A. In contrast, treatment with iExo and iMSCs resulted in significantly fewer TUNEL-positive cells than the I/R group, although the I/R + iExo and I/R + iMSC groups did not differ significantly (11.6 ± 2.8 and 16.0 ± 3.8 vs. 43.1 ± 8.3, p < 0.05 vs. the I/R group) (Fig. 3A and 3B).
Using qRT-PCR analysis, we found that iExo and iMSC treatments reduced mRNA levels of Bim and caspase-3 and restored the B-cell lymphoma 2 (Bcl-2) mRNA level compared to the I/R group (Bim, 1.92 ± 0.08 and 1.36 ± 0.11 vs. 2.61 ± 0.32; caspase-3, 1.10 ± 0.05 and 1.94 ± 0.04 vs. 1.40 ± 0.07; Bcl-2, 2.84 ± 0.09 and 2.78 ± 0.14 vs. 0.38 ± 0.05, p < 0.05 vs. the I/R group) (Fig. 3C–3E). The I/R + iMSC group demonstrated decreased Bim and caspase-3 mRNA levels compared to the I/R + iExo group.
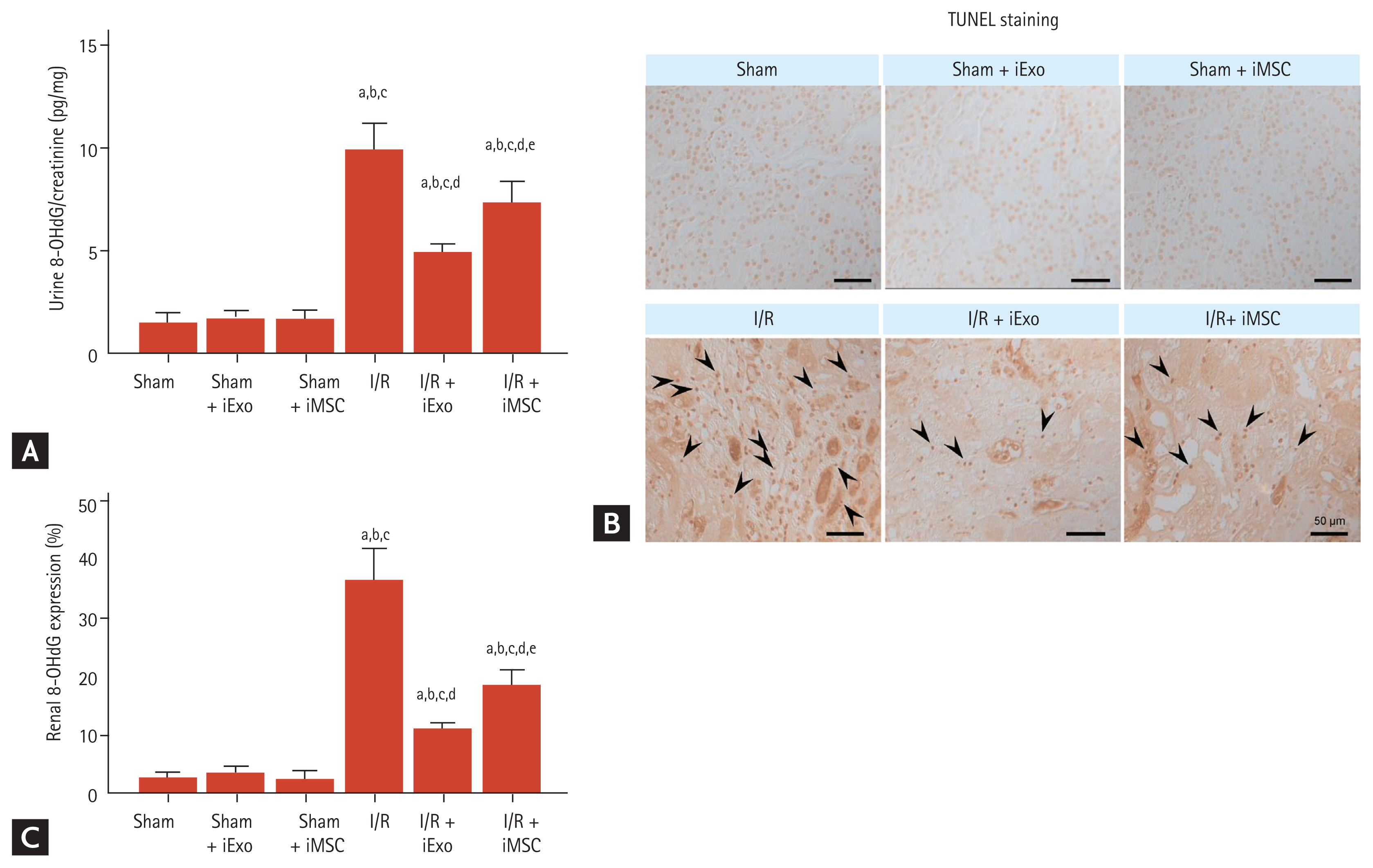
Effects of treatment with iExo or iMSCs on oxidative stress induced by renal I/R injury
I/R injury induced significantly increased urinary 8-OHdG excretion, an indicator of oxidative DNA damage in urine and kidney tissues, compared to the sham group (9.95 ± 1.2 vs. 1.41 ± 0.4, p < 0.05 vs. the sham group) (Fig. 4). However, this level was significantly decreased after treatment with iExo and iMSC (4.92 ± 0.4 and 7.35 ± 1.0, p < 0.05 vs. the I/R group). Abundant nuclear 8-OHdG immunoreactivity in kidney tissues from the I/R group was also attenuated in the I/R + iExo and I/R + iMSC groups (Fig. 4B). Fig. 5C illustrates the significantly decreased renal 8-OHdG staining after iExo and iMSC treatment during I/R (11.10 ± 0.7 and 18.57 ± 2.5 vs. 36.50 ± 5.4, p < 0.05 vs. the I/R group).
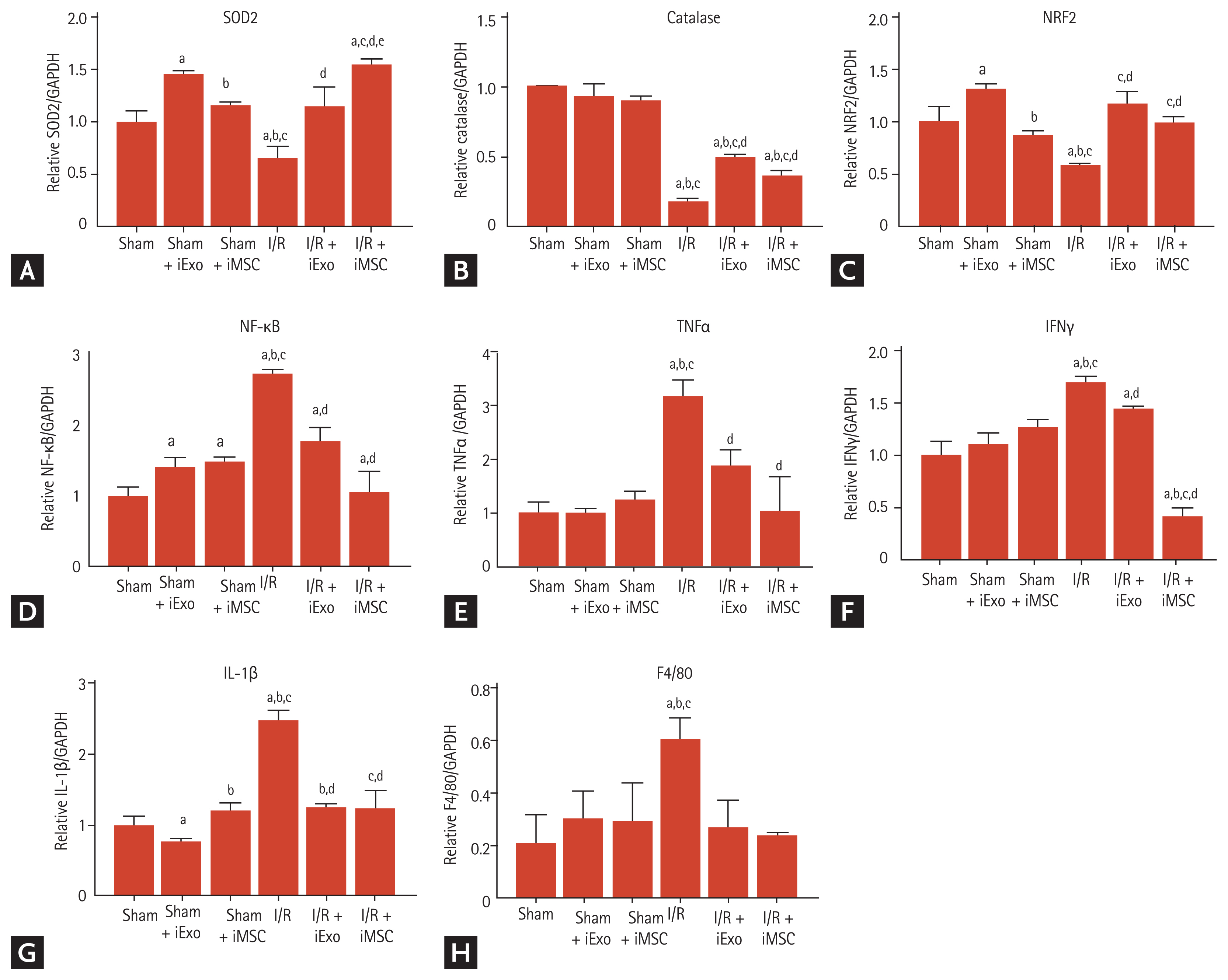
Effect of iExo or iMSC treatment on antioxidant and pro-inflammation gene expression levels
We examined the effect of treatment with iExo and iMSCs on the transcriptional levels of genes associated with antioxidants and pro-inflammation during I/R injury (Fig. 5). As expected, I/R injury significantly decreased mRNA levels of SOD2, catalase, and NRF2, which are associated with antioxidants (SOD, 0.65 ± 0.1 vs. 1.0 ± 0.1; catalase, 0.18 ± 0.01 vs. 1.0 ± 0.1, NRF2, 0.58 ± 0.1 vs. 1.0 ± 0.2, p < 0.05 vs. the sham group). However, treatment with iExo and iMSCs significantly restored these levels, as shown in Fig. 5A–5C (SOD, 1.15 ± 0.2 and 1.55 ± 0.1; catalase, 11.10 ± 0.7 and 18.57 ± 2.5; NRF2, 1.17 ± 0.1 and 0.99 ± 0.1, P < 0.05 vs. the I/R group). Consistent with these results, mRNA levels of NF-κB, TNFα, IFNγ, and IL-1β during I/R injury were also significantly attenuated by treatment with iExo or iMSC (NF-κB, 1.77 ± 0.2 and 1.06 ± 0.3 vs. 2.70 ± 0.1; TNFα, 1.87 ± 0.3 and 1.0 ± 0.7 vs. 3.20 ± 0.3; IFNγ, 1.46 ± 0.1 and 0.41 ± 0.1 vs. 1.71 ± 0.1; IL-1β, 1.25 ± 0.1 and 1.22 ± 0.2 vs. 2.47 ± 0.1, p < 0.05 vs. the I/R group).
Effects of iExo or iMSC treatment on ERK1/2 signaling pathway activation
Next, we evaluated whether the protective effects of iExo and iMSCs were associated with the signaling cascade of the extracellular signal-regulated kinase (ERK) signaling pathway, which plays an important role in cell death and survival during renal I/R injury. Activation of ERK1/2 signaling cascades was examined using immunoblot analysis based on the relative amounts of p-MEK1/2 and p-ERK1/2 (Fig. 6). Renal I/R for 25 minutes significantly decreased the ratios of both p-MEK1/2 to total-MEK1/2 (t-MEK1/2) and p-ERK1/2 to t-ERK1/2 (p-MEK1/2/t-MER1/2, 49 ± 2 vs. 100 ± 4; p-ERK1/2/t-ERK1/2, 51 ± 4 vs. 100 ± 5, p < 0.05 vs. the sham group). However, iExo or iMSC treatment significantly restored these levels (p-MEK1/2/t-MER1/2, 81 ± 2 and 73 ± 4; p-ERK1/2/t-ERK1/2, 81 ± 5 and 84 ± 4, p < 0.05 vs. the I/R group), as shown in Fig. 6. Protein expression between the I/R + iExo and I/R + iMSC groups did not differ significantly.
DISCUSSION
The results of the present study demonstrated that iExo administration could reduce renal I/R injury in a mouse model. We found that treatment with iExo attenuated renal dysfunction and the histological changes accompanying reduced inflammation, oxidative stress, and apoptosis. In addition, activation of the ERK1/2 phosphorylation signaling pathway was increased after iExo treatment during I/R injury. Therefore, we suggest that treatment with iExo effectively ameliorates renal damage following I/R injury by enhancing the cell growth and survival signaling pathway.
Many studies have shown that exosomes derived from BMSCs can alleviate tissue damage and promote functional recovery after ischemic injury [17,19]. Because iMSCs have been demonstrated to have a strong therapeutic function, we proposed that iMSC-derived exosomes may also exert powerful attenuating effects on I/R injuries. Therefore, we performed in vivo experiments using an I/R mouse model to prove our hypothesis. Consistent with our hypothesis, mice in the iExo-treated group exhibited lower sCr levels and ATN scores than those in the I/R group. Further, treatment with iExo reduced the expression of inflammatory cytokines and markers for oxidative stress and apoptosis. These data confirmed that iExo was functionally active and effective in reducing kidney damage induced by I/R.
ERK1/2 is required to repair renal tubular epithelial cells and inhibit fibrosis caused by renal injury [25], suggesting that the ERK1/2 signaling pathway is related to the pathophysiology of various renal diseases, including renal I/R [26]. In a recent study, the injection of exosomes from human-bone-marrow-derived MSC (hBMSC-Exos) into mice suffering from I/R injury resulted in improved functional recovery, histologic protection, and reduced apoptotic cell death [27]. For direct evidence, they treated human kidney 2 (HK-2) cells exposed to hypoxia/reoxygenation with hBMSC-Exos and found significantly reduced apoptosis accompanied by activation of the ERK signaling pathway in the HK-2 cells compared to the controls [27]. In this study, the I/R injury group demonstrated significant inactivation of the ERK signaling pathway with decreased p-MEK1/2 and p-ERK1/2 levels in kidney tissues compared to the sham group. However, iExo treatment significantly restored these levels, demonstrating similar results as in the iMSC treatment group. Although not explainable in this study, the exosome-enriched fraction has been reported to exhibit the highest renal protection activity from injury [28]. ERK1/2 signaling was shown to be activated in human dermal fibroblasts following treatment with exosomes collected from the culture supernatant of BMSCs [29]. Taken together, our results suggest that iExo treatment protects against I/R-induced renal injury, in which the ERK1/2 signaling pathway played an important role.
The efficacy of many MSC-based therapies has been attributed to the paracrine secretion of trophic factors, and exosomes may play a major role in mediating tissue repair [30,31]. Exosomes derived from different stem cells, such as BMSCs and adipose-derived MSCs, have been shown to attenuate injury [32–36]. In the present study, we selected exosomes derived iMSC (iExo) to treat renal I/R injury for several reasons. First, the effect of iPSC-derived MSC exosomes on renal IR injury has not been investigated. Second, iPSCs can generate unlimited amounts of early passage MSC with consistent quality. Third, iExo are a promising cell source for autologous cell therapies in regenerative medicine. Fourth, compared to adult MSCs, human iMSCs have been demonstrated to be superior regarding cell proliferation, immunomodulation, cytokine profile, generation of exosomes capable of modulating the microenvironment, and bioactive paracrine factor secretion [9,10,37].
Several studies have examined in vivo tracking to determine exosome biodistribution upon systemic delivery [38,39]. In an AKI mouse model, intravenously injected DiD-labeled extracellular vesicles (EVs), including exosomes, accumulated predominantly in the kidney of mice with AKI compared to healthy control [40]. Eventually, exosomes internalized by renal epithelial cell membrane fusion or endocytosis play a protective role in kidney injury and promote tissue regeneration and repair [41]. Until now, it is unclear how long exosomes exist in kidney lesions. However, several studies showed that exosomes have a short half-life in vivo [42,43], limiting their therapeutic potential for clinical application. Various forms of hydrogels such as chitosan/silk hydrogel, chitosan hydrogel, and imine crosslinking hydrogel have been used to enhance the therapeutic effects of exosomes [44–47]. Finally, secreted EVs can be eliminated as cellular waste. This might be a more efficient strategy for eliminating senescent proteins compared to proteasomal and lysosomal degradation [48].
Our study had several limitations. First, although the present study results are promising, the study period was only 24 hours; the long-term effects of iExo therapy for I/R injury remain uncertain. Second, although we systemically injected iExo via the tail vein, we only focused on its effects on the kidneys and not on other organs. Third, we did not observe a direct role of iExo in vitro to help elucidate its protective mechanism. Future studies are warranted to define the exact role of iExo in hypoxia-related conditions.
In conclusion, the present study demonstrated that exosomes from iPSC-derived MSC therapy effectively preserve renal function and the integrity of the kidney against I/R injury. These findings suggest a potential therapeutic approach for various types of AKI.
KEY MESSAGE
1. Mesenchymal stem cells (iMSC)-derived exosomes ameliorate renal dysfunction and the histological changes accompanying reduced inflammation, oxidative stress, and apoptosis following ischemia reperfusion injury in a mouse model.
2. iMSC-derived exosomes protect against ischemia reperfusion-induced renal injury, in which the extracellular signal-regulated kinase (ERK) 1/2 signaling pathway plays an important role.
3. Our findings suggest that iMSC-derived exosomes are useful for autologous treatment with unlimited amounts of iMSC. Also, the safety issue of cell-based therapy can be excluded.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print





