


INTRODUCTION
Although the liver has a considerable inherent regenerative capacity [1], sustained and chronic injury results in the onset of liver fibrosis. Stimuli such as viral hepatitis, alcohol, drugs, metabolic diseases, and autoimmune attack by hepatic cells trigger hepatocyte apoptosis, the impairment of the endothelial barrier, the recruitment of inflammatory cells and the activation of hepatic stellate cells (HSCs) [2-9]. Liver fibrosis is the result of an imbalance in extracellular matrix (ECM) synthesis and degradation mediated by portal fibroblasts, bone marrow-derived fibroblasts, mesenchymal cells, and activated HSCs [10]. Currently, liver transplantation is the only effective treatment for end-stage liver fibrosis [11].
Recently, stem cell transplantation has been suggested as an effective alternative therapy for hepatic diseases [12]. Alison et al. [13] and Theise et al. [14] have reported the presence of Y chromosome-positive hepatocytes in autopsied women who had received therapeutic bone marrow transplantations from male donors, suggesting the existence of pluripotent stem cells among their bone marrow cells. Moreover, stem cells, including embryonic, induced pluripotent, hematopoietic and mesenchymal stem cells (MSCs), can be differentiated into hepatocyte-like cells both in vitro and in vivo [15-17]. Of these stem cell types, MSCs have several advantages, such as easy acquisition, strong proliferative capacities and ex vivo expansion. In addition, MSCs have immune-modulatory properties and are able to migrate to damaged tissues. MSCs also secrete trophic factors, including growth factors and cytokines, which promote the regeneration of impaired tissues, including the liver.
In this review, we summarize (1) the properties of MSCs for regenerative medicine, (2) the therapeutic mechanisms of MSCs in the treatment of liver fibrosis, and (3) the clinical application of MSCs for the treatment of liver fibrosis. We also present several outstanding risks associated with their use, including their fibrogenic, tumor cell growth promotion and oncogenic potentials.
PROPERTIES OF MSCs FOR REGENERATIVE MEDICINE
MSCs are a promising source for cell-based tissue engineering and regenerative medicine. MSC transplantation is considered safe and has been widely tested in clinical trials of cardiovascular, neurological and immunological diseases with encouraging results. The properties of MSCs can be represented by their basic characteristics as stem cells and their therapeutic potentials as drugs. With regard to their basic characteristics, MSCs have the potential for self-renewal and differentiation into multiple types of cells. Sufficient numbers of these MSCs can be expanded without the loss of their potential for clinical application. In addition, MSCs can move toward areas of injury in response to signals of cellular damage, which are known as homing signals. This migration property of MSCs is important in regenerative medicine because various injection routes can be used depending on the damaged tissue or organ. MSCs can be transplanted into the liver by intravenous, intraperitoneal, intrahepatic, intrasplenic, or portal-venous injection, although the reported effectiveness has differed slightly based on the injection route and research group. MSCs are characterized by low expression of human leukocyte antigen (HLA) class I molecules and the absence of major histocompatibility complex (MHC) class II antigens, Fas ligand and the co-stimulatory molecules B7-1, mB7-2, CD40, and CD40L. These reduced immunogenic expression profiles cause MSCs to have immuno-tolerant phenotypes, allowing them to be used in allogeneic transplantation [18,19].
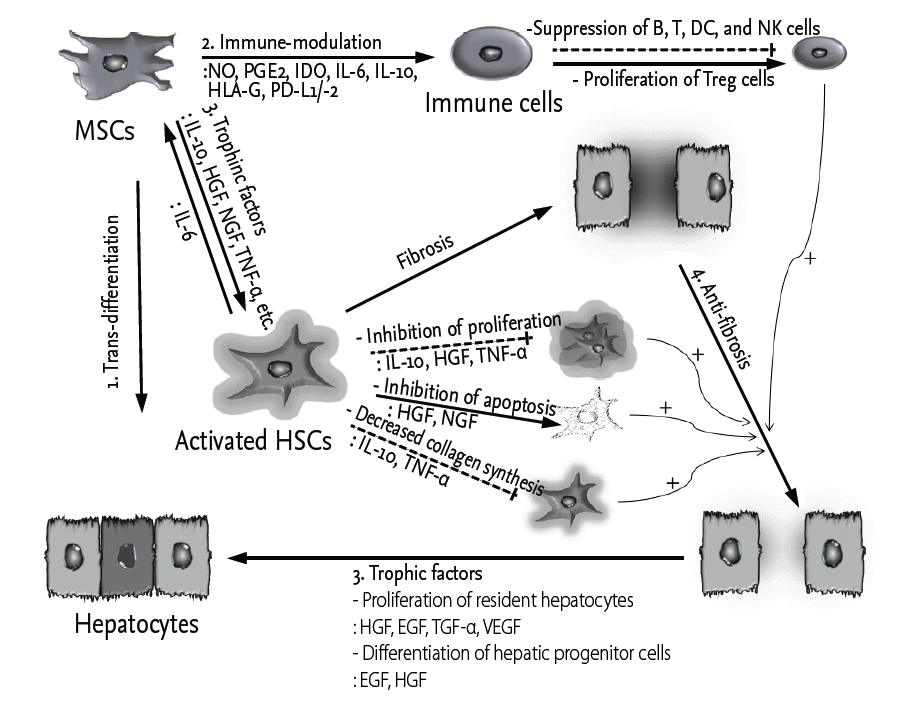
The therapeutic properties of MSCs that are relevant to liver fibrosis are related to their capacities for hepatocyte-like differentiation and their immune-modulatory, trophic factor secretory, anti-fibrotic, and anti-oxidant activities (Fig. 1). MSCs can be differentiated into multiple cell lineages, including hepatocytes, both in vivo and in vitro. They play immune-modulatory roles both in the adaptive and innate immune systems by suppressing T- [20], B- [21,22], dendritic [23,24], and nature killer (NK) cells [25] or promoting the generation of regulatory T (Treg) cells via a mechanism involving interleukin 10 (IL-10) [26,27]. MSCs express various trophic factors (growth factors and cytokines) that stimulate resident cells and matrix remodeling to promote the differentiation of native progenitor cells and the recovery of injured cells. In fibrotic tissue, MSCs can down-regulate myofibroblasts and lead to anti-fibrotic activity. In addition, MSCs display cytoprotective effects by inducing anti-oxidant response elements (AREs) in carbon tetrachloride (CCl4)- and thioacetamide (TAA)-induced liver injury [28,29].
THERAPEUTIC MECHANISMS OF MSCs IN THE TREATMENT OF LIVER FIBROSIS
Hepatocyte-like cell differentiation
Hepatocyte-like cells differentiated from MSCs are promising sources of liver regeneration. MSCs can be differentiated into hepatocyte-like cells by treatment with hepatocyte growth factor (HGF), fibroblast growth factor (FGF)-2/-4, epidermal growth factor (EGF), oncostatin M, leukemia inhibitory factor, dexamethasone, insulin-transferrin-selenium, and/or nicotinamide [30]. Hepatocyte-like cell differentiation can also be induced by co-culture with liver cells [31] and pellet culture [32]. Moreover, human bone marrow-derived MSCs can be differentiated into hepatocyte-like cells without fusion in allyl alcohol-treated rat livers [33]. However, the trans-differentiation of MSCs into hepatocytes has been rarely observed (less than 1% of the total liver mass) in animal models in relation to the amounts injected [34].
Immune-modulatory potential of MSCs
MSCs can express various soluble factors, such as nitric oxide, prostaglandin E2 (PGE2), indoleamine 2,3-dioxygenase (IDO), IL-6, IL-10, and HLA-G. These soluble factors regulate the proliferation and functions of a variety of immune cells and induce Treg cells [35]. In particular, PGE2 increases the anti-inflammatory cytokine IL-10 and decreases tumor necrosis factor α (TNF-α), interferon γ (IFN-γ), and IL-12 in dendritic cells (DCs). PGE2 also reduces IFN-γ and IL-4 in Th1 and Th2 cells and stimulates the proliferation of Treg cells [35]. Moreover, IDO and HLA-G suppress the proliferation of effector T cells, inhibit the maturation of DCs, inhibit the proliferation and immunoglobulin G secretion of B cells and reduce the cytotoxicity of NK cells [36,37]. In addition to the secretion of soluble factors from MSCs, these cells can suppress the activation of immune cells through direct cell-cell contact. MSCs can inhibit T-cell proliferation by inducing the apoptosis of effector T cells by promoting the association of programmed death-1 (PD-1) with its ligands PD-L1 and PD-L2, and MSCs are capable of rendering T cells anergic by down-regulating the expression of the co-stimulatory molecules CD80 and CD86 on antigen-presenting cells [38-40]. Unbalanced immune cell populations or immune cell infiltration of the liver can disrupt its immune-privileged state, resulting in liver injury or fibrosis. Therefore, the immune-modulatory potential of MSCs plays an important role in the treatment of liver fibrosis.
Secretion of trophic factors by MSCs
Accumulating evidence has revealed that various trophic factors secreted by MSCs play key therapeutic roles in regenerative medicine. MSCs express trophic factors, such as growth factors, cytokines, and chemokines, which are known not only to reduce the inflammation, apoptosis and fibrosis of damaged tissues but also to stimulate angiogenesis and tissue cell regeneration [41]. Moreover, after MSCs move to damaged sites for repair, they are stimulated by local factors, such as inflammatory cytokines, ligands of Toll-like receptors and hypoxic conditions. These stimuli lead to the production of a large amount of growth factors that perform multiple functions to achieve tissue regeneration [42,43]. Hepatocytes in fibrotic livers reach replicative senescence after many rounds of injury and repair; however, trophic factors secreted by MSCs can lead to the survival of living and dying hepatocytes via anti-apoptotic (stromal cell-derived factor 1, HGF, insulin-like growth factor 1 [IGF-1], and vascular endothelial growth factor [VEGF]), mitogenic (EGF, HGF, nerve growth factor [NGF], and transforming growth factor α [TGF-α]), and angiogenic effects (VEGF) [44-46]. Anti-apoptotic events that correlate with reduced inflammation have been observed in fibrotic tissues following MSC transplantation in association with alterations in HGF and IGF-1 expression [47,48]. HGF, EGF, and TGF-α, which are potent mitogens, are primarily associated with hepatocyte proliferation [49-52], and VEGF enhances angiogenesis, which is responsible for liver regeneration. In addition to hepatocytes, hepatic progenitor cells, which are located in the canals of Hering, can be differentiated into hepatocytes or biliary lineage cells following treatment with EGF or HGF, respectively [53]. Trophic factors, such as IL-10, HGF, NGF, TGF-β, and TNF-α, regulate the proliferation of activated HSCs and decrease collagen synthesis in liver fibrosis.
Anti-fibrotic activities of MSCs
Liver fibrosis, which is the precursor to cirrhosis, is the result of the deposition of ECM proteins and is mediated primarily by activated HSCs. Following liver injury, HSCs undergo a phenotypic switch from quiescent, vitamin A-storing cells to proliferative, α-smooth muscle actin (SMA)-positive, myofibroblast-like cells with increased collagen synthesis. Interestingly, the anti-fibrotic activities of MSCs have been reported in various fibrotic animal models in the heart, liver, kidneys, lungs, peritoneum, pancreas, skin, and rectum [54]. MSCs suppress the pathophysiological process that is mediated by chronic inflammation, and this immunosuppressive mechanism contributes to a modification of the microenvironment; the result is diminished tissue fibrosis, increased resident stem cell proliferation and eventually tissue regeneration.
Moreover, MSCs are able to reduce the proliferation of activated HSCs and collagen synthesis through indirect or direct cell-cell contact. In indirect contact mode, trophic factors (i.e., IL-10, HGF, TGF-β3, and TNF-α) that are secreted by MSCs inhibit the proliferation of HSCs and decrease collagen synthesis [55,56], while HGF and NGF promote the apoptosis of HSCs [55,57]. MSCs that are directly co-cultured with HSCs significantly suppress the proliferation and α-SMA expression of HSCs through cell-cell contact, and this activity is partially mediated by Notch pathway activation [58]. Furthermore, MSCs can regulate the expression of matrix metalloproteinases (MMPs) and tissue inhibitors of MMPs (TIMPs). In several fibrosis models, MSCs have been shown to increase the expression of MMPs (i.e., MMP-2, -9, -13, and -14) [59-61] or to decrease TIMP-1 expression [60,62], and these alterations are generally associated with fibrosis resolution.
Anti-oxidant activities of MSCs
Reactive oxygen species (ROS) trigger oxidative stress, which induces liver diseases such as liver fibrosis, cirrhosis, viral hepatitis, hepatocellular carcinoma (HCC), and others [63-67]. CCl4 and TAA are toxins used worldwide to generate experimental liver injury [29,68,69]. These toxins stimulate ROS production, which results in hepatocyte damage through lipid peroxidation and the alkylation of proteins, nucleic acids, and lipids [29,70-72]. MSCs have been shown to overcome CCL4- and TAA-induced oxidative stress in vitro and to reduce liver injury through anti-oxidant activities in vivo [28,29]. The up-regulation of ROS in CCl4-treated liver cells has been reported to be attenuated by co-culturing with MSCs via an increase in superoxide dismutase activity and the induction of AREs, which represents a cytoprotective response in the injured liver [29]. Additionally, MSCs protect hepatocytes by reducing ROS damage that is induced by TAA both in vivo and in vitro [28].
CLINICAL APPLICATION OF MSCs FOR LIVER FIBROSIS
Clinical trials using MSCs have been designed to investigate their therapeutic potentials for the treatment of cirrhosis (Table 1). In a phase 1 trial, autologous bone marrow-derived MSCs were infused through the peripheral veins of four patients with decompensated cirrhosis. There were no side effects reported in these patients during follow-up, and the Mayo End-Stage Liver Disease score was improved in half of the patients. Furthermore, the qualities of life of all four patients improved by the end of follow-up [73]. In a phase 1-2 trial, Kharaziha et al. [74] showed an improvement in liver function in cirrhosis patients who were injected with 30 to 50 million autologous MSCs via the peripheral or portal veins. In phase 2 trials [75-80], Jang and colleagues [79] showed the beneficial effects of autologous bone marrow MSC transplantation for the treatment of alcoholic cirrhosis. MSCs (5 × 107 cells) were injected into the hepatic artery twice at weeks 4 and 8. According to the Laennec fibrosis system, histological improvement was observed in 6 of 11 patients (54.5%). The Child-Pugh score was improved in ten patients (90.9%), and the levels of TGF-β1, type 1 collagen, and α-SMA significantly decreased after MSC therapy [79]. Similar results were obtained by Amer et al. [75], Amin et al. [76], Zhang et al. [77], El-Ansary et al. [78], Peng et al. [80] (Table 1). Autologous bone marrow-derived hepatocyte-like cells also improved the liver function of 20 patients with endstage liver failure when these cells were transplanted via intrasplenic and intrahepatic routes [75]. In addition to the improvement of liver function by MSCs, the incidence of HCC or mortality in patients with hepatitis B-related liver failure showed no significant difference between autologous MSCs-injected and control groups after 192 weeks of follow-up [80]. In addition to autologous bone marrow-derived MSCs, allogeneic MSCs from umbilical cords have been used to improve liver function in decompensated cirrhosis patients [77].
Although MSCs have been widely used in clinical and pre-clinical studies of liver fibrosis, several issues must be carefully considered, including their fibrogenic potentials and capacities for tumor cell growth promotion and oncogenicity. Depending on the MSC injection route and liver disease status, MSCs can differentiate into myofibroblasts rather than hepatocytes [81,82]. Engraftment of human MSCs is very low in normal and acutely injured livers compared to chronically injured livers. Moreover, a significant number of human MSCs exhibit a myofibroblast-like morphology during acute liver injury [81]. Baertschiger et al. [82] observed that stable engraftment of MSCs in the liver is not achieved following intrasplenic injection; however, after intrahepatic injection, MSCs permanently remain in the liver but primarily differentiate into myofibroblasts. Another risk of MSC transplantation has been identified with regard to the susceptibility of these cells to malignant transformation and the promotion of pre-existing tumor growth. As mentioned above, MSCs can secrete various growth factors (i.e., FGFs, EGF, TGF-β, HGF, and VEGF) that promote tumor cell growth and neo-vascularization [83]. Although the malignant transformation of human MSCs has not been reported in clinical trials, the risk of the introduction of genetic mutations during the ex vivo expansion of MSCs must be carefully considered prior to transplantation. Despite these risks, the therapeutic effects of MSCs in liver fibrosis have been verified in pre-clinical and clinical studies. Thus, these cells can be expected to become a new treatment for liver fibrosis in the near future.
FUTURE PROSPECTS OF MSC THERAPY FOR LIVER FIBROSIS
To develop MSC therapy for liver fibrosis, larger clinical studies must be conducted to obtain meaningful insights into the safety and clinical efficacy of MSC infusion [84,85]. Accumulating evidence has revealed that autologous MSC infusion is safe because autologous MSCs do not induce an immune reaction; in particular, autologous MSCs expand the Treg population and reduce the T cell population [86]. Reports of the efficacy of MSCs are controversial and depend on the research group; however, we expect that issues regarding their efficacy will be resolved in the near future by a large multicenter randomized clinical trial that is currently being conducted in Korea. Moreover, multicenter international clinical studies on the safety and efficacy of MSC treatments for liver fibrosis can help clinicians reach a consensus on the treatment of liver fibrosis, which will ultimately improve the prognosis of patients.
Several important considerations must still be addressed to support stem cell therapies for liver fibrosis treatment. First, the delivery route of MSCs into the liver has not been standardized but is important for optimizing their therapeutic effects and engraftment. Second, the number of injections of MSCs and their concentrations must be optimized to improve therapeutic effects. Third, the survival duration of engrafted MSCs is important for achieving sustained efficacy. In many pre-clinical animal studies, human MSCs have been observed by immunohistochemical analysis using human-specific markers [87-89]; however, more sophisticated techniques to identify and follow the fates of injected MSCs will be required for clinical translation. For instance, MSCs can be labeled with superparamagnetic iron oxide nanoparticles or reporter genes, causing them to be traceable using advanced imaging technologies [90-95]. Because nanoparticles or reporter genes can modify the properties of MSCs, biomarkers specific to injected MSCs that do not cause cell damage must be developed, even if developing such tools takes a long time.
CONCLUSIONS
MSCs are potentially relevant therapeutic agents for the treatment of liver diseases because of their potential to differentiate into hepatocytes as well as their immune-modulatory properties and ability to secrete trophic factors. Nevertheless, MSC therapy needs to be further evaluated in large randomized and controlled clinical trials with longer follow-up periods. In addition, further studies are needed to solve various issues, including those involving the fibrogenic potential of MSCs and their ability to promote pre-existing tumor cell growth.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print





