


INTRODUCTION
Asthma is associated with an enhanced Th2 immune response1). Th1 and Th2 cells are regulated reciprocally, and it has been shown that Th1 enhancers such as bacille Calmette-Gu├®rin vaccine2) and CpG oligodeoxynucleotides3) suppress asthmatic reactions in animal models. The adrenal androgen dehydroepiandrosterone (DHEA) is also reported to increase the production of Th1 cytokines, including interleukin (IL)-24, 5) and interferon (IFN)-╬│5), and to prevent or attenuate allergic inflammatory airway responses in an animal model of asthma6, 7).
In a study on human peripheral blood mononuclear cells (PBMCs), the number of cells secreting IFN-╬│ was related with the serum level of DHEA sulfate ester (DHEAS)8). In addition, patients with asthma9, 10) or atopic dermatitis11), another Th2-associated disease, showed significantly lower levels of serum DHEA compared with healthy controls. Moreover, Tabata et al.11) showed that DHEA significantly reduced IL-4 production in PBMCs from patients with atopic dermatitis.
The purpose of this study was to examine whether DHEA suppresses Th2 cytokine production in cultured PBMCs from asthmatic patients.
MATERIALS AND METHODS
Subjects
Sixty-one consecutive suspected asthmatic or non-asthmatic men (age range, 18~34 years) were recruited from Chonnam National University Hospital. Most of the patients with suspected asthma were young men who required a medical certificate for asthma before they could be exempted from obligatory military service. Most non-asthmatic subjects were medical students and doctors. Female subjects were also recruited, but only 18 subjects responded (age range, 21~38 years). Participants were classified according to gender and the presence of airway hyperresponsiveness (AHR), based on their responses to a methacholine bronchoprovocation test. All controller medications for asthma, including oral steroids, were withheld for more than 1 week before enrollment. The Institutional Review Board of Chonnam National University Hospital approved this study. All subjects were informed of the experimental procedures and provided written informed consent.
Study design
On the day of the study, a medical history was obtained and a physical examination was performed. Allergy skin-prick tests, baseline spirometry, and a methacholine bronchoprovocation test to detect AHR were conducted, in addition to several laboratory analyses, including complete blood counts, total IgE levels, and quantification of serum DHEAS and cortisol. A blood sample was drawn for PBMC culture. The fresh blood was immediately processed to isolate PBMCs, which were cultured with and without DHEA. The culture supernatant was assayed for Th1 and Th2 cytokines via an enzyme-linked immunosorbent assay (ELISA).
Asthma tests
Allergy skin-prick tests were carried out using the following 10 common aeroallergen extracts (Allergopharma, Reinbek, Germany): Dermatophagoides farinae, D. pteronyssinus, cockroach, cat, dog, Aspergillus fumigatus, hazel, birch, timothy, and ragweed. Histamine (1 mg/mL) and saline (0.9%) solutions were used as positive and negative controls, respectively. Reactivity was graded on a scale of 0~4+ for each allergen, based on the ratio of the size of the allergen-induced wheal to the size of the histamine-induced wheal. The sum of all 10 grades was used as an overall "atopy score," and the sum of the scores for D. farinae and D. pteronyssinus was designated the "house dust mite score." Total serum IgE levels were measured by nephelometry (Behring Diagnostics GmbH, Germany).
The bronchial challenge tests followed a standardized tidal breathing method as described previously12). Aerosols were generated using a DeVilbiss 646 nebulizer (DeVilbiss Co., Somerset, PA, USA; output, 0.13 mL/min), and airway responses to methacholine (Sigma, St. Louis, MO, USA) aerosols were measured using a spirometer (Spiro Analyzer ST-250, Fukuda Sangyo, Tokyo, Japan). The provocation concentration of methacholine (PC20) that resulted in a 20% decrease in forced expiratory volume in 1 s (FEV1) was calculated by linear interpolation of the log dose-response curve. AHR was defined as methacholine PC20 < 16 mg/mL. An index of bronchial reactivity (BRindex) was calculated as log10 [10 + the maximal% fall in FEV1/log10 (dose in mg/dl of the stimulus required to produce maximal% fall)]13).
Hormone and cytokine assay
Serum concentrations of DHEAS and cortisol were measured via radioimmunoassay using commercially available kits (Diagnostic Products Corp., Los Angeles, CA, USA). Radioactivity was quantified using a gamma counter (Cobra 5010 Quantum; Packard Instrument Co., Meriden, CT, USA).
PBMCs were isolated by centrifugation over a Ficoll-Hypaque density gradient (Lymphoprep; Nycomed Pharma, Oslo, Norway). The PBMCs were washed five times and re- suspended at a concentration of 1├Ś106 cells/mL in RPMI medium 1640 (Gibco BRL, Paisley, UK) supplemented with 10% fetal bovine serum (FBS; Gibco, Grand Island, NY, USA) and 1% penicillin/streptomycin/amphotericin B (BioWhittaker, Walkersville, MD, USA). The PBMCs were stimulated with a T-cell-specific mitogen, concanavalin A (Con A; 2.5 ┬Ąg/mL; Sigma), after pre-incubation with DHEA (10 ┬Ąmole; Sigma) for 1 h at 37Ōäā. Some PBMCs were also pre-incubated with 0.01~1 ┬Ąmole DHEA. After 48 h of culture, cell culture supernatants were collected and stored at 70Ōäā until analysis. The concentrations of IFN-╬│, IL-5, and IL-10 were measured using commercially available ELISA kits (Biosource, Camarillo, CA, USA). Assay sensitivities for IFN-╬│, IL-5, and IL-10 were 4, 4, and 1 pg/mL, respectively.
Statistical analysis
Data are presented as the mean┬▒standard error of the mean (SEM). PC20 values and IgE, hormone, and cytokine levels were log-transformed before analysis. The intra-group difference in cytokine levels was determined using the Wilcoxon signed rank and Friedman tests. The Mann-Whitney U test was used to detect significant differences between groups. Associations between variables were examined using Spearman's rank correlation coefficient. A value of p<0.05 was considered statistically significant.
RESULTS
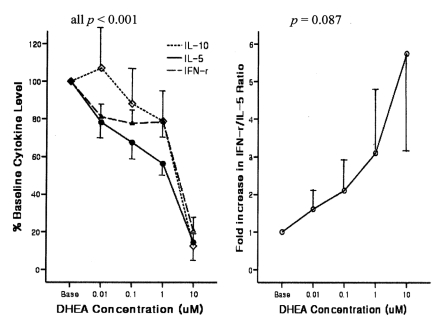
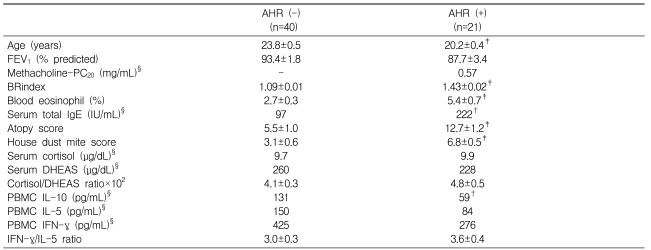
Male subjects with AHR were younger and showed significantly higher BRindices, eosinophil proportions, total IgE levels, atopy scores, and house dust mite scores, but lower IL-10 levels compared with men without AHR (Table 1). In subjects with AHR, the addition of DHEA to Con A-stimulated PBMCs significantly suppressed the production of IL-10, IL-5, and IFN-╬│ in a dose-dependent manner (all p<0.001) and tended to increase the IFN-╬│/IL-5 ratio (p=0.087; Figure 1). In subjects without AHR, DHEA also significantly suppressed the production of IL-10, IL-5, and IFN-╬│ in a dose-dependent manner (all p<0.001) but did not affect the IFN-╬│/IL-5 ratio.
The observed changes in cytokine levels and the IFN-╬│/IL-5 ratio from baseline after treatment with the highest dose (10 ┬ĄM) of DHEA were significant in all comparisons, except for changes in the IFN-╬│/IL-5 ratio in men with AHR, which showed a trend toward increasing (p=0.088) (Table 2). The maximal dose of DHEA markedly suppressed IL-5 production in subjects with AHR (24.0┬▒7.8%) compared with those without AHR (40.9┬▒3.6%, p<0.01). The suppression of IFN-╬│ production was also significantly greater in subjects with AHR (29.7┬▒7.0%) than in those without AHR 54.5┬▒5.1%, p<0.01). Although the resulting increase in the IFN-╬│/IL-5 ratio tended to be higher in subjects with AHR compared with those without AHR, the difference between these groups did not reach statistical significance (p=0.71).
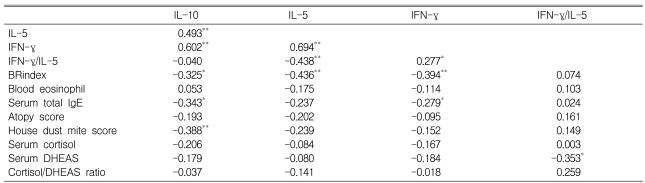
In male subjects, significant positive relationships were detected among IL-10, IL-5, and IFN-╬│ in terms of the degree of DHEA (10 ┬ĄM)-induced suppression (expressed as % of baseline value; Table 3). The suppressive effect of DHEA on IL-10 was significantly related to AHR, serum total IgE level, and reactivity to house dust mites. In addition, both IL-5 and IFN-╬│ suppression were significantly related to AHR, IFN-╬│ suppression was significantly related to serum total IgE concentrations, and the IFN-╬│/IL-5 ratio was significantly related to serum DHEAS.
Compared with women without AHR (n=15), men without AHR (n=40) showed a significantly higher level of DHEAS (geometric mean: 153 vs. 260 ┬Ąg/dL, ; p<0.01) but not cortisol (8.2 vs. 9.7 ┬Ąg/dL), which resulted in a significantly lower cortisol/DHEAS ratio (6.1┬▒0.9 vs. 4.1┬▒0.3%, p<0.05). Compared with female subjects, male subjects showed significantly higher eosinophil proportions, total IgE levels, and atopy scores, but lower IL-10 levels (199 vs. 131 pg/mL, p<0.01). Similar to male subjects, female subjects with AHR exhibited trends toward cytokine suppression (IL-10: 13.3┬▒1.9%, IL-5: 44.2┬▒7.0%, IFN-╬│: 68.4┬▒7.4%; all p<0.01) and a Th1 shift (╬öIFN-╬│/IL-5 ratio: 2.5┬▒0.7; p<0.01). However, we were unable to evaluate the effect of DHEA on cytokine production in female subjects with AHR owing to the small sample size (n=3), despite the similar trends in the suppression and Th1 shift.
DISCUSSION
In this study, DHEA suppressed the production of IL-5 in cultured PBMCs, resulting in a non-significant increase in the IFN-╬│/IL-5 ratio in subjects with AHR. It has been reported that DHEA suppresses allergic inflammatory airway responses6, 7) and serum concentrations of IgE and IL-614) in animal models of asthma, and that DHEA reduces IL-4 production in cultured PBMCs from patients with atopic dermatitis11). The mechanism by which DHEA suppresses allergic responses is thought to involve a shift in the Th1/Th2 balance toward Th1. Our results support this hypothesis.
The Th1 cytokine IFN-╬│ inhibits Th2 cells15) and plays an important role in regulating the allergic airway reaction16, 17). However, in contrast to some reports that DHEA increases IFN-╬│ production5, 18), we found that DHEA suppressed IFN-╬│ production in a dose-dependent manner. Similar results have been reported by Yu et al.6, 7) and Araneo et al.19). Therefore, our results suggest that a mechanism mediated by some other factor (i.e., not IFN-╬│) mediates the DHEA-associated suppression of the Th2 cytokine IL-5. Unfortunately, we did not quantify the number of PBMCs in our study; however, it has been reported that DHEA suppresses the proliferation of Con A-stimulated splenocytes in mice20). In addition, DHEA antagonizes glucocorticoid-induced deviation of the immune response toward a Th2 cytokine profile, probably through the peroxisome proliferator-activated receptor ╬▒21). Alternatively, DHEA may down-regulate both IFN-╬│ and IL-5 production while maintaining a Th1 bias, which would suggest that the balance between Th1 and Th2 is more important than the actual level of Th1 cytokines in determining the influence of DHEA on asthma.
IL-10 drives the generation of T regulatory 1 cells22), which produce IL-10, and in turn, IL-10 inhibits both Th1 and Th2 cytokine production23). Asthma is associated with diminished IL-10 expression in the allergic airway24), which is consistent with our result showing that the baseline IL-10 levels were lower in male subjects with AHR than in those without AHR. However, the IL-10 concentration in PBMCs was significantly reduced by treatment with DHEA, and we observed a significant positive correlation between the changes in IL-10 and IL-5 levels. Yu et al.6) also showed that DHEA decreased the production of both IL-5 and IL-10. Although it has been demonstrated that DHEA induces IL-10 production in spleen cells25), the decreased production of IL-10 may be the result of suppressed cell proliferation20) and other mechanisms.
Relatively more severe atopy or asthma was associated with a greater degree of DHEA-induced cytokine suppression in this study. Because inflammatory/immune cells are activated during active allergic disease processes and because DHEA up-regulates DHEA receptor activity in activated human T lymphocytes 26), DHEA would likely be more effective in suppressing inflammatory/immune responses during active allergic diseases.
As DHEA is an adrenal androgen, it is expected that the serum DHEAS level and cortisol/DHEAS ratio would differ significantly between male and female subjects without AHR. It has been reported that IFN-╬│ production in PBMCs is positively correlated with the plasma DHEA level and inversely correlated with the cortisol/DHEA ratio27). Therefore, the effects of DHEA treatment are likely influenced by gender. In the present study, exogenous DHEA increased the IFN-╬│/IL-5 ratio in an inverse relationship with the serum DHEAS level, even though cytokine suppression and increased IFN-╬│/IL-5 ratios tended to occur in both genders. These results are consistent with our previous study suggesting that in men, the higher background levels of DHEA and other androgens associated with the Th1 bias may result in an adaption to DHEA, and thus greater Th1-inducing power may be necessary to overcome this adaptation in men28).
The number of female subjects with AHR in this study was too small to evaluate the effects of DHEA among AHR subjects and between AHR and non-AHR subjects. Thus, further investigation involving a much larger sample size is required. Furthermore, because serum concentrations of DHEAS and cortisol are influenced by many factors, precise measurement is required in this regard.
Collectively, our study demonstrated that DHEA decreased both IL-5 and IFN-╬│ production in cultured PBMCs and skewed the Th1/Th2 balance toward Th1. In addition, the suppressive effect of DHEA on cytokine production was related to the severity of AHR or atopy. Therefore, DHEA supplementation may have therapeutic benefits for asthmatics.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print





