


INTRODUCTION
The first identifiable marker in the development of an antibody-producing plasma cell from the bone marrow precursor cell is the rearrangement of the immunoglobulin heavy-chain genes which is followed by the rearrangement of light chain genes1).
At this point the stem cell becomes the pre-B cell which contains heavy chains in its cytoplasm. The pre-B cell gives rise to B-cells bearing their surface immunoglobulin and other markers1,2). Antigens binding to immunoglobulin on B-cell surface initiate the B-cell proliferation and terminal differentiation into antibody-producing plasma cells.
This process of B-cell maturation is also brought about by Epstein-Barr virus3,4) and other plant and bacterial products known as poly-clononal B-cell activators.
Many of them require T-helper cells and macrophages for B-cells to induce maximum immunoglobulin response.
In the studies of human immunoglobulin synthesis, polyclonal activiators pokeweed mitogen or Epstein-Barr virus have been extensively utilized5).
These studies could define stages of B-lymphocyte differentiation and the roles played by networks of immunoregulatory T-cells and macrophages in the B-cell maturation.
These studies clarified the pathogenic mechanisms of primary immunodeficiency disease6) autoimmune disorders7,8) and acquired immunodeficiency disorders9,10).
Recently the effects of interferons have been extensively studied and interferon is being introduced as a theurapeutic agent for various disorders in the clinic11,12).
In addition to their antiviral effects13,11) interferons are reported to have a variety of immunologic functions, such as inhibition of cellular proliferation13ŌĆō16) activiation of the natural killer cells17ŌĆō19) or the enhancement of the cytotoxicity of the immune T-cells20).
Interferon is also known to have modulatory activities on B-lymphocyte activiation21ŌĆō23). Inteferon suppressed the antibody response to antigen when interferon was given to cells 6 hour prior to or up to 48 hours after antigen challange21). However, pretreatment of B-cells with low amounts of Interferon or late addition of interferon to antigen stimulated culture led to enhanced antibody synthesis.
Gisler et al21) found inteferon did not affect T-helper function or macrophage function in their system and concluded Interferon acted directly on B-cells. Choi, et al24) studied the effects of Interferons on human B-cell differentiation with the polyclonal activator pokeweed mitogen.
They found that the effect of interferon was dose-dependent; enhancing B-cell differentiation at low dose, while suppressing it at high dose.
Their experiments suggested the increased production of immunoglobulin was due to macrophage activation by interferon and the suppression of immunogobulin production was the result of impaired T-helper cell proliferation.
At a high dose of interferon, B-cell proliferation was directly suppressed.
Harfast et al25) also found that depending on the time of exposure, interferon either enhanced or suppressed the pokeweed mitogen-induced immunoglobulin synthesis by human peripheral blood lymphocytes. They and Pelton and Denman26) found that interferon acted exclusively on B-cells.
The aims of the present study were as follows.
First, to further investigate the effects of interferons on B-cell differentiation induced by polyclonal activators.
Using T-dependent (pokeweed mitogen) and T-independent mitogen (Epstein-Barr virus) and other techniques, the mechanism of interferon action on the B-cell differentiation in vitro was studied.
Second, to verify the effect of the conventional virus-induced interferon preparations as the real actions of inteferon.
By comparing the effect of conventional inteferon preparations with the effect of recombinant forms of interferon this study could exclude the effects of possible impurities in the former preparations.
Additionaly, the relative potency of different types of inteferons (IFN-╬▒ and IFN-╬│) was studied.
MATERIALS AND METHODS
1. Preparation of Cells
Peripheral blood was obtained from normal adult volunteers.
Mononuclear cells were Isolated by Ficoll-Hypaque density gradient centrifugation. T-and B-enriched cell fractions were obtained by rosetting with neuraminidase-treated sheep erythrocytes27).
Mononuclear cells and sheep erythrocytes were mixed in a ratio of 1:50, centrifuged on a Ficoll-Hypaque gradient in siliconized tubes. Cells at the interface were subjected to a second cycle of sheep erythrocytes rosetting to further separate the rosette forming cells. The interface cells obtained after 2 cycles of rosetting were obtained as B-enriched cells which contained less than 5% T-cells. The rosette forming cells obtained after hypotonic shock in 0.9% ammonium chloride solution were used as T-cells which contained less than 5% non-T. The T-and Be-cells were were washed twice and were suspended in RPMI-1640 medium plus 10% fetal calf serum containing glutamine(1:1000 v/v) and antibiotics (1: 1000 v/v) (Grand Island Biological Co. NY, USA).
2. Interferon Preparations Used
Virus-induced human leukocyte IFN-╬▒ (Polyferon, Bioferon, West Germany) and recombinant IFN-╬▒ (rIFN-╬▒) and recombinant IFN-╬│ (rIFN-╬│) (Dr. karl Thomae Ltd., west Germany) were used.
3. Induction of Ig Production
For mononuclear cell culture, 1 ├Ś106/ml cells in 12├Ś75 mm plastic tubes (Falcon, MD, USA) were stimulated with 10 ╬╝g/ml of pokeweed mitogen (Grand Island Biological Co., NY, USA).
For coculture of T-and Be-cells, 2├Ś106/ml T-cells and the same number of Be-cells were mixed together into round bottomed 96-well plates (Nunc, denmark) in a total volume of 0.2 ml. Culture were incubated for 12 days at 37┬░C in humidified air containing 5% CO2.
In experiments using Epstein-Barr virus as the polyclonal activiator of B-cells, the mononuclear cells at a concentration of 3├Ś106/ml were incubated with culture supernatants of B-95-8 cell line for 2 hrs, washed twice and plated in round-bottom culture plates at a concentration of 2├Ś106/ml in volumes of 0.2 ml. Desired amounts of rIFN-╬│ were added to cultures.
4. Determination of IgM by ELISA
An enzyme-linked immunosorbent assay (ELISA) modified from Kelly et al.28) was used. Microtiter plates (Dynatech, Va, USA) were coated with 0.2 ml of affinity column-purified rabbit anti-human IgM (Atlantic Antibodies, Me, USA). The anti-Ig had been diluted in 0.1 M sodium bicarbonate buffer, PH 9.6, to a concentration of 1 ╬╝g/ml.
The plates were incubated at 4┬░C overnight and stored at ŌłÆ20┬░C and used in less than 4 weeks. The coated plates were extensively washed with phosphate buffered saline (PBS) containing Tween 20 (0.05% v/v) (Tween-PBS).
Each well received 0.2 ml of Tween-PBS containing 5% of culture supernatant to be tested or serial dilutions of known amounts of IgM.
After 4 hrs incubation at room temperature or overnight incubation at 4┬░C, the plates were extensively washed with Tween-PBS and 0.2 ml of alkaline-phosphatase-conjugated polyvalent anti-lg in Tween-PBS (1 ╬╝g/ml) (TAGO Inc., CA, USA) were added.
After 2 hrs incubation at room temperature, the plates were again washed extensively with Tween-PBS and 0.2 ml of P-nitrophenyl phosphate disodium (1 mg/ml, Sigma, St. Loius, USA) in 0.1 M sodium bicarbonate buffer, pH 8.6, containing 0.001 M MgCl2 were added to each well.
After 1hr incubation at room temperature, the absorbance at 492 nm was measured with a Dynatech automatic reader.
In each experiment a standard curve was constructed from samples containing known amounts of IgM (Nor-Partigen, Behringwerke, West Germany).
All cultures were run in triple and each sample was measured duplicate by ELISA.
5. Preincubation of Celluar Fractions With IFN
The effect of pretreating T-cells or B-cells with IFN on PWM-induced Ig production was studied by incubating 2├Ś108/ml cells with different IFN concentrations for 16 hrs at 4┬░C.
The cells were then washed twice, resuspended in culture medium at desired cell concentration.
The IFN-treated T-cells were recombined with untreated Be-cells and the IFN-treated Be-cells were recombined with untreated T-cells.
6. Identification of T-Cell Subsets in Culture
The effect of preincubation of T-cells with IFN on T-cell subset composition was studied by testing the T-cell marker profile of the lymphocytes at the termination of the culture.
T-cell subsets were determined with the immunoenzyme technique.
Leu-4, Leu-3 and Leu-2 (Beckton-Dickinson, CA, USA) monoclonal antibodies were utilized.
RESULTS
1. Effects of IFN-╬▒ on lg Production by Lymphocytes
Virus-induced human leukocyte IFN-╬▒ added to the lymphocyte culture in a dose of 300 U/ml enhanced Ig production, most prominently when it was given 24 hrs prior to PWM stimulation (Table 1).
The effects of IFN-╬▒ at doses 10,100,1000, and 10,000 U/ml at the time of culture start, 48 hrs, 72 hrs and 96 hrs after the culture start are shown in Fig. 1. At all the doses employed, IFN-╬▒ stimulated Ig production when it was added until 48 hrs after culture start (+ 18%, +76% of control). The addition of IFN-╬▒ after 72 hrs from culture start had a suppressive effect, while at the dose of 10,000 U/ml it retained stimulatory activity.
2. Effects of Recombinant Interferon-╬│ on Ig Production by Lymphocytes
In a preliminary study, the addition of 100 U/ml of rIFN-╬│ to lymphocyte culture enhanced the Ig production three-to five-fold (Table 2).
This effect was noted when rIFN-╬│ was added to culture 24 hrs prior to, at the time of culture or 72 hrs after the culture start.
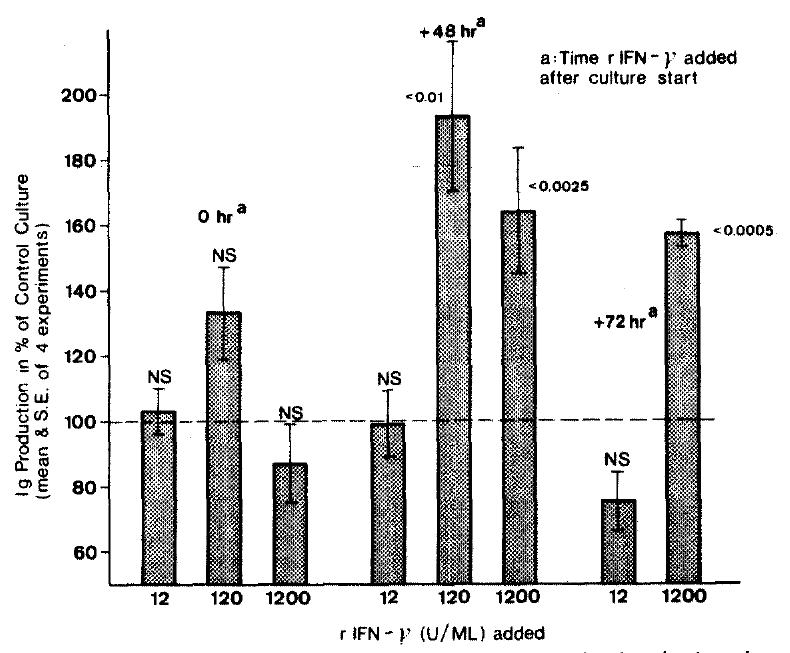
In further experiment employing doses of r-IFN-╬│ 12,120 and 1200 U/ml (Fig. 2), the dose of 120 U/ml rIFN-╬│ given 48 hrs after culture start was most stimulating. It enhanced the Ig production by 94%.
3. Effects of Preincubation of Cell Fractions with IFNs on Subsequent Ig Production
When T-cells preincubated with IFN were recombined with untreated autologous Be-cells, Ig production was enhanced (+ 85% and +487% of control) (Table 3). Conversely, when B-enriched cells were preincubated with IFNs and then recombined with untreated T-cells, there was marked suppression of subsequent Ig production (<20 ng/ml) (p<0.05).
4. Effects of Preincubation of T-Cells with IFNs on T-Cell Subset Composition at the Termination of Culture
To study the effects of IFNs on the proliferation of T-cell subsets, the T-cell subset profile was determined in the cultures at the termination of culture, on day 12 (Table 4). The cultures which contained T-cells preincubated with rlFN-╬▒ or with rIFN-╬│ had increased helper to suppressor T-cell ratio in comparison to control culture (p<0,025).
5. Effects of Recombinant Interferon-╬│ on Ig Production Induced by EBV
At a dose of 10 U/ml, rIFN-╬│ stimulated Ig production induced by EBV (+ 112%) (Table 5). At higher doses (100 U/ml and 500 U/ml), rIFN-╬│ had significant suppressive activity on EBV-induced B-cell differentiation.
DISCUSSION
In studying the regulation of Ig production in the human system, the polyclonal activator PWM has been widely used.
The merit of this system is that analysis of the respective functions of T-helper, T-suppressor, macrophage or B-cells is possible.
After the polyclonal activation, the secreted Igs can be assessed by hemolytic plaque assay or quantitiated by double antibody radioimmunoassay or Enzyme-liked immnuo-sorbent assay28).
The present study employed polyclonal activators PWM and EBV and quantitated the secreted Ig by ELISA method to study the effects of inteferons on human B-cell maturation.
By employing T-dependent system using PWM as well as the pure B-cell mitogen EBV, the cell types on which IFN acted could be dissected.
The ELISA techique is an objective and sensitive method for measuring Ig levels.
The results of the present study show that IFNs have modulatory activity on polyclonal activator (PWM or EBV)-induced Ig production by human peripheral blood lymphocytes. In accordance with previous results24,25,29), IFN enhanced or suppressed the Ig production of lymphocytes depending on time and dose of addition.
When low to moderately high dose of IFN-╬▒ (10 to 1000 U/ml) were added to culture until 48 hrs after PWM stimulation, it had stimulatory activity.
With a very high dose of IFN-╬▒ (10,000 U/ml) or with r-IFN-╬│ the Ig production was enhanced until 72 hrs after the PWM-stimulation of the culture.
The effects of virus-induced interferon and the recombinant types of interferons were similar in their actions, suggesting that the action of the former preparations were not that of possible contaminating lymphokines.
The effects of recombinant inteferon-╬│ were most striking and enhanced the Ig procuction 2-to 6-fold of control cultures. In EBV-induced Ig production by peripheral blood lymphocytes, r-IFN-╬│ displayed regulatory activities: at low dose (10 U/ml) enhancing and at higher doses (100 U/ml and 500 U/ml) suppressing Ig production. Since EBV is known to stimulate B-cells independent of accessory cells3,4) this regulatory effect of IFN on B-cell differentiation may be its direct effect on B-cells, as was reported by other group30). This modulatory effect of IFN on EBV-driven Ig production by human lymphocyte culture is first reported in this study. A novel interpretation of the regulatory effects of IFN on antibody production is that of Gresser13).
According to him, low blood level of IFN at initiation of viral infection could amplify the antibody response while high blood level of IFN at termination of viral infection could suppress the further B-cell activiation.
In co-culture studies, T-cells preincubated with IFN were recombined with untreated B-cells under PWM stimulation. This culture produced more Ig than control culture and contained more T-helper cells than in control culture, suggesting IFN preincubation of T-cells favors T-helper cell proliferation, which then stimulates B-cells to produce more Ig.
Thus the mechanism of action of IFN on in vitro Ig production by human lymphocytes seems two-fold; direct regulatory action on B-cell activation as reported by Gisler et al21) Harfast et al25) and Pelton and Denman26) as well as through T-helper cell mediation as was reported by Sonnenfeld et al29).
To sum up above results; 1) A low dose of IFN stimulates B-cell differentiation in vitro.
2) A moderately high dose of IFN stimulates T-helper cell proliferation which then sitimulates B-cell differentiation.
3) B-lymphocytes exposed to moderately high to high dose of IFN were directly suppressed in differentiation.
Recently, using anti-immunoglobulin (anti-╬╝) or Staphylococcus aureus Cowan strain (SAC) for B-cell activation followed by addition of factors for maturation of B-lymphocytes, more direct analysis of B-cell differentiation events became possible31,32). SAC activted B-cells were induced to proliferate by interleukin-2 but secreted no Ig. The subsequent addition of interferon-╬│ brought Ig secretion30). Sidman et al23) also found that IFN is one of B-cell differentiation factors. As clinical application of IFN is being carried out, it may be interesting to study the effects of in vivo application of IFN on the regulation of Ig production.
Another subject for further study would be the effects of various lymphokines on normal and malignant B-cell proliferation and differentiation after anti-╬╝ or SAC induced activation.
CONCLUSION
In accordance with the previous results24,25) the present study showed that IFN has a modulatory activity on polyclonal activator-induced Ig production by human peripheral blood lymphocytes: depending on the time and dose of addition, IFN either enhanced or suppressed the Ig production by MNCs.
This modulatory effect of IFN has also been demonstrated in the EBV-driven Ig production by human MNCs, which is first reported in this study.
In coculture studies, T- or B-cells were preincubated with IFNs and recombined with untreated autologous B- or T-cells for culture under PWM stimulation, preincubation of B-cells with moderately high dose of IFN prior to PWM-stmulation with T-cells suppressed the subsequent Ig production.
On the other hand, T-cells preincubated with IFN prior to PWM-stimulation with B-cells produced increased amount of Ig. The T-cell subset analysis at the termination of these culture showed enhanced proliferation of T-helper cells relative to T-suppressor cells.
Thus the mechanism of action of IFN on in vitro Ig production by lymphocytes seems twofold; direct regulatory action on B-cell activation as well as through T-helper cell mediation.
Compared to IFN-╬▒, IFN-╬│ showed 2- to 6-fold stimulating activity on Ig production by MNCs. The present study employed the recombinant forms of interferon as well as virus-induced leukocyte interferon to compare their effects on in vitro B-cell differentiation. The effects of both preparations of interferons were similar on B-cell differentiation.
This result confirms the previous experiments which used the conventional leukocyte-IFN preparations.
Further studies on the effects of interferons on the diffferentiation of malignant B-cell disorders would be meaningful.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print





