


INTRODUCTION
In general, when an organism is placed under sudden stress, the level of activity of the cells exciting complement of enzyme increases, leading to a spurt in production. Epinephrine synthesis in the adrenal medulla is also subject to the biochemical influence of .the adrenal cortex. Because blood from the cortex passes through the medulla, most of the chromaffin cells are exposed to high levels of cortical steroids, When the steroids are absent, it has been found that the enzyme that catalyzes the conversion of norepinephrine into epinphrine is degraded at an unusually high rate, leading to the decline of its cellular level. Thus, the intimate anatomical relation of the cortex and the medulla extends to the interaction at molecular level as well.
Stress is known to induce the secretion of hormones from the adrenal cortex as well as from the meulla. The cortical hormones apparently ensure that the synthesis of epinephrine is maintained1). Kohler and his collaborators2) found that estradiol and progesterone administration to intact female rats increased adrenal tyrosine hydrosylase activity. Moreover, tyrosine hydroxylase activity is known to be increased significantly in diethylstilbesterol-induced pituitay tumors of the rats than in control pituitaries. These data support the possible existence of local catecholaminergic mechanism(s) that could be modulating prolactin secretion from pituitary tumors and from ectopic pituitaries3).
In hypothalamic and vocal control nuclei of male zebra finches, all hormone-induced changes in noradrenergic neurotrasmission are estrogen-dependent, while modulation of dopaminergic function is much more variable and frequently androgen-dependent4). Steroids are also known to affect catecholamine (CA) synthesis5), degradation6), release7) and re-uptake9).
However, the steroid-induced hyperpolarization in guinea-pig ganglion neurons in vitro was accompanied by a change in the input resistance of the cell, indicating an involvement of some kind(s) of ion channel(s) in the action of glucocorticoid10). It has been also shown that brief hyper-polarization and increased potassium conductance are produced by 17-╬▓-estradiol in rat medial amygdala brain slices11). Moreover, according to the results of Miguel and his co-workers12). estradiol seems to decrease the ability of the adrenal medulla to release CA to the peripheral blood, whereas progesterone most affects tyrosine hydrosylase activity. Being that its effects temporally and partially depend on estrogens.
It has been also found that 17-╬▒-estradiol inhibits DMPP- and high K+-induced CA release from the cat adrenal glands and 45Ca++ uptake into bovine chromaffin cells in culture stiulated by DMPP or high K+13). More recently. Laret and his collaborators14) have suggested that the amygdaloid catecholaminergic systems exert an inhibitory effect on CA content of the adrenals and the ovary, and influence the ovarian estradiol secretion mechanism. This influnce of the amygdala in the regulation of the ovary could be mediated by an efferent pathway from the central nervous system to the ovary.
Thus, there are many controversial reports on CA release from the adrenal medulla. The present study was designed to examine the effect of 17-╬▒-estradiol on CA secretion from the isolated perfused rat adrenal glands and to clarify the mechanism of its action.
MATERIALS AND METHODS
1. Experimental Procedure
Male Sprague-Dawley rats, weighing 180 to 250 grams, were anesthetized with ether. The adrenal gland was isolated by the method described previously15). The abdomen was opened by a midline incision and the left adrenal gland and surrounding area were exposed by placing three hook retractors. The stomach, intestine and portion of the liver were not removed, but pushed over to the right side and covered by saline-soaked gauge pads, and urine In the bladder was removed in order to obtain enough working space for tying blood vessels and cannulations.
A cannula, used for perfusion of the adrenal gland(Fig. 1-A), was inserted into the distal end of the renal vein after all branches of the adrenal vein(if any), vena cava and aorta, were ligated. Heparin (400 IU/ml) was injected into vena cava to prevent blood coagulation before ligating vessels and cannulations. A small slit was made into the adrenal cortex just opposite the entrance of the adrenal vein. Perfusion of the gland was started, making sure that no leakage was present, and the perfusion fluid escaped only from the slit made in the adrenal cortex. Then the adrenal gland, along with ligated blood vessels and the cannula, was carefully removed from the animal and placed on a platform of a leucite chamber. The chamber was continuously circulated with water heated at 37┬▒1 ┬░C (Fig. 1-B)
2. Perfusion of Adrenal Gland
The adrenal glands were perfused by means of a ISCO pump (WIZ Co.) at a rate of 0.3 ml/min. The perfusion was carried out with Krebs-bicarbonate solution of following composition (mM):NaCl, 118.4:KCl, 4.7:CaCl2, 2.5:MgCl2, 1.18:NaHCO3, 25:KH2PO4, 1.2:glucose, 11.7. The solution was constantly bubbled with 95% O2┬▒5% CO2 and the final pH of the solution was maintained at 7.4┬▒0.05. The solution contained disodium EDTA (10 ug/ml) and ascorbic acid (100 ug/ml) to prevent oxidation of catecholamine.
3. Drug Administration
The perfusions of DMPP (100 uM) and McN-A-343 (100 uM) for 2 minutes and/or a single injection of ACh (5.32 mM) and KCl (56 mM) in a volume of 0.05ml were made into the perfusion stream via a three way stopcock, and Bay-K-8644 (10-5M) was also perfused for 4 min.
In the preliminary experiments it was found that, upon administration of the above drugs, secretory responses to ACh, KCl, McN-A-343 and Bay-K-8644 returned to pre-injection level in about 4 min, but the responses to DMPP in 8 min.
4. Collection of Perfusate
As a rule, prior to each stimulation with cholinergic agonists or excess K+, perfusate was collected for 4 min to determine the spontaneous secretion of CA (background sample). Immediately after the collection of the background sample, collection of the perfusates was continued in another tube as soon as the perfusion medium containing the stimulatory agent reached the adrenal gland. Stimulated samples were collected for 4 to 8 min. The amounts secreted in the background sample have been subtracted from those secreted from the stimulated sample to obtain the net secretion value of CA, which is shown in all of the figures.
To study the effects of 17-╬▒-estradiol and its antagonist on the spontaneous and evoked secretion, the adrenal gland was perfused with Krebs solution containing 17-╬▒-estradiol for 20 min. the perfusate was collected for a certain background sample, and then the medium was changed to the one containing the stimulating agent and the perfusates were collected for the same period as that for the background sample. Generally, the adrenal glandŌĆÖs perfusate was collected in chilled tubes.
5. Measurement of Catecholamines
CA content of perfusate was measured directly by the flurometric method of Anton and Sayre16) without the intermediate purification alumina for the reasons described earlier15) using fluorospectrophotometer (Schimadzu Co. Japan).
A volume of 0.2 ml of the perfusate was used for the reaction. The CA content in the perfusate of stimulated glands by secretogogues used in the present work was high enough to obtain readings several fold greater than the reading of control samples (unstimulated). The sample blanks were also lowest for perfusates of stimulated and non-stimulated samples. The content of CA in the perfusate was expressed in terms of norepinephrine (base) equivalents.
6. Statistical Analysis
All data are presented as means with their standard errors, and the significance of differences were analyzed by StudentŌĆÖs paired t-test using the computer system as previously described17).
7. Drugs and Their Sources
The following drugs were used: 17-╬▒-estradiol, 17-╬▓-estradiol, acetylcholine chloride, 1.1-dimethyl-4-phenyl piperazinium iodide(DMPP), norepinephrine bitartrate. methyl-1, 4-dihydro-2, 6-dimethyl-3-nitro-4-(2-trifluoro-methylphenyl)-pyridine-5-carboxylate (BAY-K-8644), metoclopramide hydrochloride (Sigma Chemical Co., U.S.A.). tamoxifen citrate, (3-(m-cholro-phenyl-carbamoyl-oxy)-2butynyl trimethyl ammonium chloride [McN -A-343] (RBI, U.S.A). Drugs were dissolved in distilled water (stock) and added to the normal Krebs solution as required, except Bay-K-8644, 17-╬▒-estradiol, 17-╬▓-estradiol and tamoxifen, which were dissolved in 99.5% ethanol and diluted appropriately (final concentration of alcohol was less than 0.1%). Concentrations of all drugs used are expressed in terms of molar base.
RESULTS
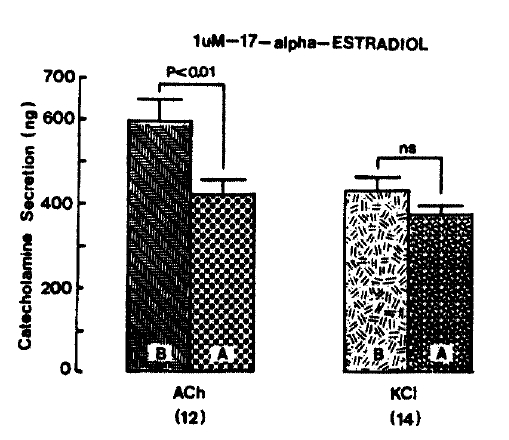
1. Effect of 1 uM 17-╬▒-estradiol on CA Secretion Evoked by ACh, Excess K+, DMPP and McN-A-343 from the Perfused Rat Adrenal Glands
After the initial perfusion with oxygenated Krebs-bicarbonate solution for 1 hr, basal CA release from the isolated perfused rat adrenal glands amounted to 24.3┬▒ 2.8 ng/2 min (n = 8). Being a typical and widely used female sexual hormone, it was decided initially to examine the effects of 17-╬▒-estradiol on cholinergic receptor stimulatin, as well as membrane depolarization-mediated CA secretion from perfused rat adrenal glands. Secretogogues were given at 20 to 30 min-intervals. 17-╬▒-estradiol was present 20 min before each stimulation. In the present study, it was found that 17-╬▒-estradiol itself did not produce any effect on basal CA output (data not shown).
When ACh (5.32 mM), in a volume of 0.05 ml. was injected into the perfusion stream, the amounts of CA secreted was 595.0┬▒53.5 ng for 4 min. However, after the pre-perfusion with 1 uM 17-╬▒-estradiol for 20 min, Ach-stimulated CA sectrtion was significantly decreased to 421.3┬▒ 32.5 ng (p<0.01) for 4 min from 12 adrenal glands as shown in Fig. 2. On the other hand, it has been found that depolarizing agent like KCI sharply stimulates CA sectretion. In the present work, excess K+ (56 mM)-stimulated CA secretion after the pre-treatment with 1 uM 17-╬▒-estradiol for 20 min was not affected. In the presence of 1 uM 17-╬▒-estradiol, it amounted to 372.9┬▒21.8 ng (ns) for 4 min as compared with its corresponding control secretion of 431.8┬▒ 31.4 ng for 4 min from 14 glands (Fig. 2).
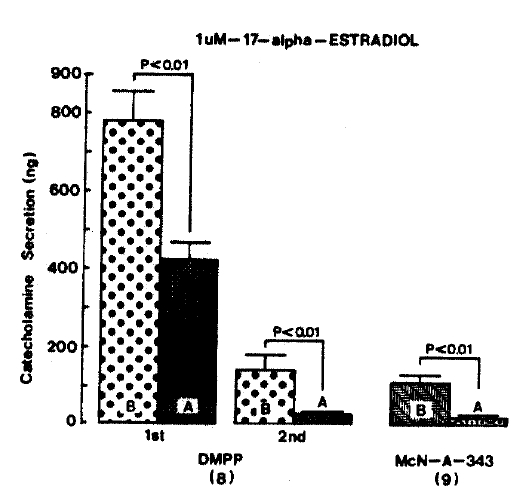
When perfused through the rat adrenal gland, DMPP (100 uM for 2 min), which is a selective nicotinic receptor agonist in autonomic sympathetic ganglia, evoked a sharp and rapid increase in CA secretion. As shown in Fig. 3, DMPP-stimulated CA secretion before pre-loading with 1 uM 17-╬▒-estradiol was 781.9┬▒77.2 ng (0ŌĆō4 min) and 147.9┬▒32.8 ng (4ŌĆō8 min), while after pre-treatment with 1 uM 17-╬▒-estradiol for 20 min they were greatly reduced to 425.6┬▒45.4 ng (0ŌĆō4 min, p<0.01) and 30.0┬▒8.7 ng (4ŌĆō8 min, p<0.01), respectively from 8 rat adrenal glands. As illustrated in Fig. 3. McN-A-343 (100 uM), which is a selective muscarinic Mi-agonist18), perfused into and adrenal gland for 2 min caused an increased CA secretion to 113.3┬▒24.5 ng for 4 min from 9 experiments. However, McN-A-343-stimulated CA secretion in the presence of 1 uM 17-╬▒-estradiol was markedly inhibited to 28.3┬▒4.6 ng (p<0.01) for 4 min, which is 25% of the corresponding control secretion.
2. Effect of 10 uM 17-╬▒-estradiol on CA Secretion Evoked by ACh, Excess K+, DMPP, McN-A-343 and Bay-K-8644 from the Perfused Rat Adrenal Glands
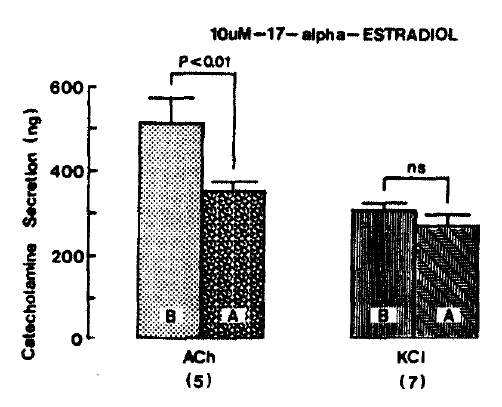
In order to test the dose-dependent effects of 17-╬▒-estradiol on cholinergic receptor-stimulated CA secretion, as well as membrane depolarization-mediated secretion, more increased concentration of 17-╬▒-estradiol to 10 uM was pre-loaded into the adrenal medulla. Fig. 4 shows that 10 uM 17-╬▒-estradiol-pre-treatment greatly exerts inhibition of CA secretion evoked by ACh. but not by excess KCI. In the present study, ACh (5.32 mM)-stimulated CA secretion prior to pre-loading with 10 uM 17-╬▒-estradiol was 513.0 ┬▒ 56.6 ng for 4 min from 5 rats. However, under 10 uM 17-╬▒-estradiol effect, which was perfused 20 min before stimulation was induced, it was markedly inhibited to 351.0 ┬▒ 26.2 ng (p<0.01) for 4 min, which was 68% of its corresponding control secretion. Excess K+ (56 mM)-stimulated CA secretion in the presence of 10 uM 17-╬▒-estradiol amounted to 269.3┬▒25.3 ng/4 min from 7 glands, as compared to the corresponding control secretion of 308.6┬▒12.2 ng/ 4 min. There was no significant difference between the both groups of before and after pre-treatment with 10 uM 17-╬▒-estradiol, as shown in Fig. 4.
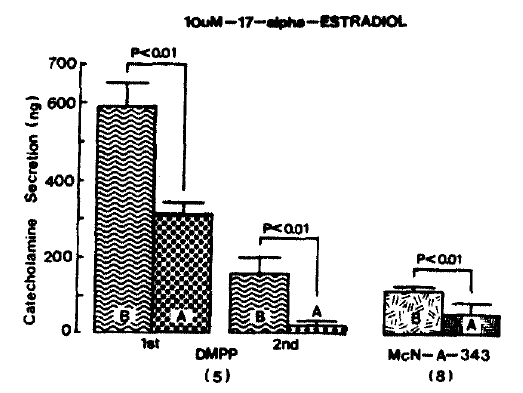
Nicotinic receptor agonist. DMPP (100 uM) perfused into the adrenal gland evoked great CA sectrtion of 592.5 ┬▒ 57.5 ng (0ŌĆō4 min) and 160.0┬▒44.3 ng (4ŌĆō8 min). while following perfusion with 10 uM 17-╬▒-estradiol for 20 min they were markedly reduced to 312.5 ┬▒ 27.2 ng(0ŌĆō4 min, p<0.01) and 27.5 ┬▒ 7.2 ng(4ŌĆō8 min, p<0.05) from 5 adrenal glands as compared with their control responses, respectively as shown in Fig. 5.
In 5 rat adrenal glands, McN-A-343 (100 uM)-stimulated CA sectretion was 114.0 ┬▒ 12.7 ng/4 min before administration of 10 uM 17-╬▒-estradiol but in the presence of 10 uM 17-╬▒-estradiol, McN-A-343-evoked CA secretion was significantly decreased to 57.0 ┬▒ 27.0 ng (p<0.01) of its control secretion as shown in Fig. 5.
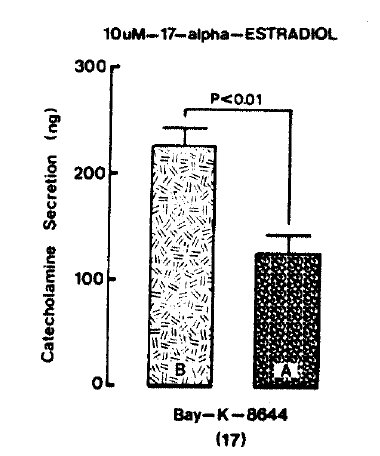
Since Bay-K-8644 is known to be as a calcium channel activator and to cause positive inotropy and vasconstriction in isolated tissues and intact animals19,20) and to enhance basal Ca++ uptake21) and CA release22), it was of interest to determine the effects of 17-╬▒-estradiol on Bay-K-8644-stimulated CA secretion from the isolated perfused rat adrenal glands. Fig. 6 illustrates the inhibitory effect of 10 uM 17-╬▒-estradiol on Bay-K-8644-evoked CA secretion. Bay-K-8644 (10 uM), given into the perfusion stream for 4 min, increased CA secretion to 224.5┬▒18.8 ng from 17 rat adrenal glands. However, under the effect of 10 uM 17-╬▒-estradiol which was pre-loaded 20 min before Bay-K-8644 was introduced. Bay-K-8644-stimulated CA secretion was strikingly depressed to 126.1┬▒17.3 ng (p<0.01) for 4 min as compared to the corresponding control releases. Thus, the release was reduced to 56% of the control secretion.
3. Effect of 100 uM 17-╬▒-estradiol on CA Secretion Evoked by ACh, Excess K+, DMPP McN-A-343 from the Perfused Rat Adrenal Glands
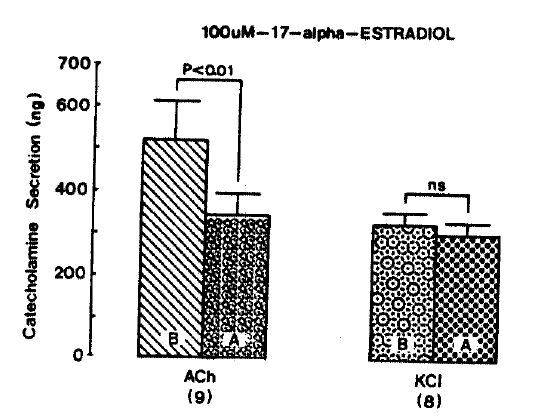
It was tried to examine the effects of 17-╬▒-estradiol, as a maximal concentration in the present experiment, on cholinergic receptor-stimulated as well as membrane depolarization-mediated CA secretion from the isolated perfused rat adrenal glands. Prior to pre-loading with 100 uM 17-╬▒-estradiol, CA secretion evoked by a single injection of ACh (5.32 mM) and excess K+ (56 mM) in a volume of 0.05 ml into an adrenal gland was 521.7┬▒87.4 ng and 331.9┬▒22.9 ng for 4 min. respectively as shown as in Fig. 7. However, following the pre-loading with 100 uM 17-╬▒-estradiol for 20 min, ACh-induced CA release was greatly blocked to 345.0┬▒ 50.9 ng (p<0.01, n = 9) for 4 min while excess K+-induced release was 308.3┬▒30.4 ng (ns, n = 8), which was not modified as compared to its corresponding control secretion (Fig. 7).
Fig. 8 shows the blockade of 100 uM 17-╬▒-estradiol to CA secretory effect evoked by DMPP and McN-A-343 from the rat adrenal glands. In the present work, in the absence of 100 uM 17-╬▒-estradiol, DMPP (100 uM)-and McN-A-343 (100 uM)-evoked CA secretion amounted to 747.5┬▒97.6 ng(0ŌĆō4 min) and 198.0┬▒52.3 ng (4ŌĆō8 min), and 169.3┬▒32.0 ng (0ŌĆō4 min), respectively, while in the presence of 100 uM 17-╬▒-estradiol. which was pre-loaded 20 min before stimulation, they were prominently depressed to 362.5┬▒44.9 ng (0ŌĆō4 min, p<0.01, n = 6) and 12.0┬▒4.8 ng (4ŌĆō8 min, p<0.01, n = 6) and 38.6┬▒6.4 ng (0ŌĆō4 min, p<0.01, n = 7), respectively.
4. Effect of 17-╬▓-estradiol on CA Secretion Evoked by ACh, Excess K+, DMPP and McN-A-343 from the Isolated Rat Adrenal Glands
In the previous experimental results, as shown in Fig. 3ŌĆō8, it was found that 17-╬▒-estradiol showed a dose-dependent inhibition in CA secretory responses. Moreover, It has been known that administration of 17-╬▓-estradiol to 3-week ovariectomized fish (Clarias batrachus) results in both seasonal and differential effects on the CA levels. After 3-weeks of ovariectomy, CA levels were elevated significantly in the preparatory, pre-spawning and spawning phases, and were unaltered in the post-spawning season23). Therefore, it is likely of interest to examine the effect of 17-╬▓-estradiol on CA secretion evoked by various secretagogues.
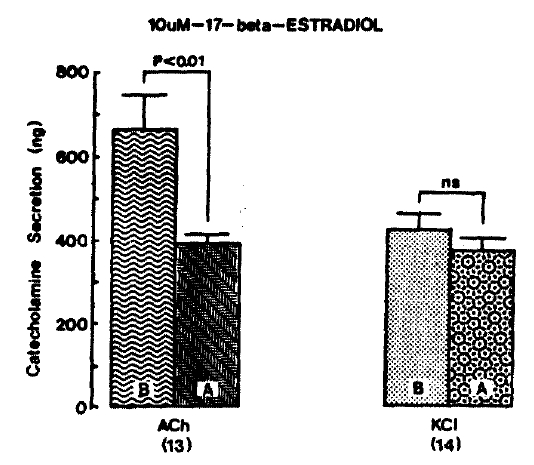
CA release evoked by ACh (5.32 mM) and excess K+ (56mM) after pre-loading with 10 uM 17-╬▓-estradiol for 20 min amounted to 390.0┬▒28.2 ng (p<0.01, n=13) and 372.9┬▒32.7 ng (ng. n = 14) for 4 min, respectively, as compared to each corresponding control secretion of 662.3┬▒82.0 ng and 428.2┬▒32.3 ng for 4 min as shown in Fig. 9.
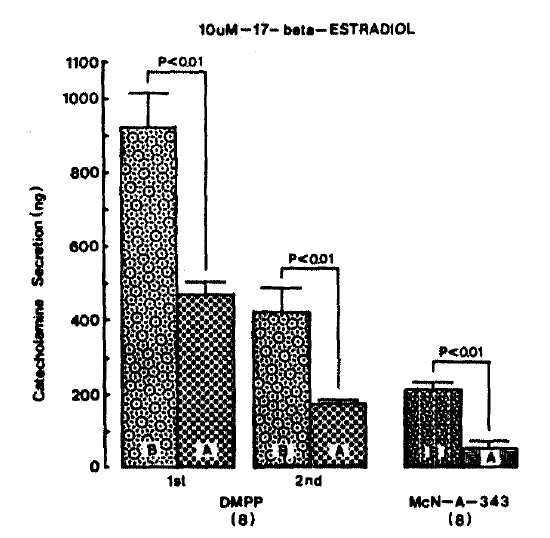
DMPP (100 uM)-and McN-A-343 (100 uM)-stimulated CA releases in the absence of 17-╬▓-estradiol were 920.6┬▒96.5 ng (0ŌĆō4 min) and 425.6┬▒63.6 ng (4ŌĆō8 min), and 219.4┬▒17.1 ng (0ŌĆō4 min), respectively. However, after pre-loading with 10 um 17-╬▓-estradiol for 20 min, they were significantly reduced to 470.6┬▒35.6 ng (0ŌĆō4 min, p<0.01, n = 8) and 176.3┬▒8.9 ng (4ŌĆō8 min, p<0.01, n = 8), and 56.3┬▒16.0 (0ŌĆō4 min, p<0.01, n = 8), respectively, as compared to their corresponding control secretion. Fig. 10 illustrates that 17-╬▓-estradiol markedly inhibits CA release evoked by DMPP and McN-A-343.
5. The Effects of 17-╬▒-estradiol Plus Tamoxifen on CA Release Evoked by ACh, Excess K+, DMPP and McN-A-343
Since it has been found that tamoxifen is a competitive inhibitor of estrogen binding at the estrogen receptor24ŌĆō26), it was tried to determine the effect of 17-╬▒-estradiol in the presence of tamoxifen on CA secretion evoked by various secrettagogues from the isolated rat adrenal glands.
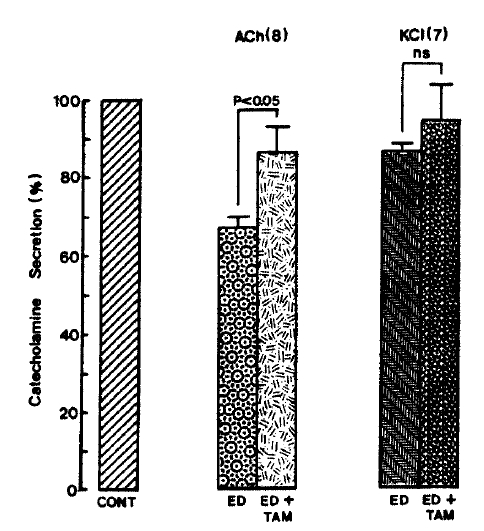
When given into an adrenal vein in a volume of 0.05ml. ACh (5.32 mM)-and excess K+(56mM)-induced CA releases, in the presence of 10uM 17-╬▒-estradiol along with 2 uM tamoxifen for 20 min, were considerably recovered to 86.6┬▒6.3% (p<0.05, n=8) and 94.3┬▒9.3% (ns, n=7) of their control secretion (100%), respectively, as compared to their secretory responses of 67.4┬▒2.3 and 86.7┬▒1.9% of their controls in the presence of 17-╬▒-estradiol only(Fig. 10).
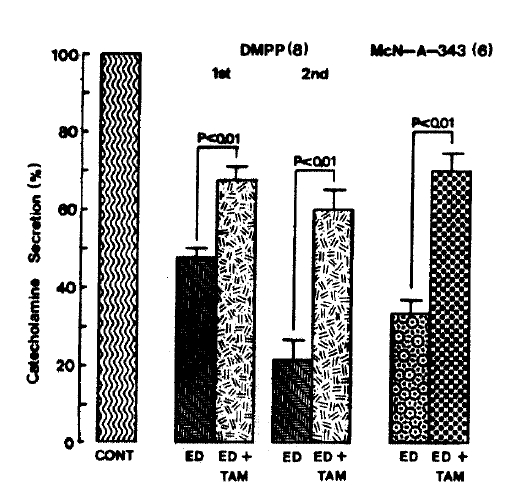
On the other hand. 17-╬▒-estradiol plus tamoxifen treatment did fail to alter the basal CA secretory response (data not shown). As depicted in Fig. 12. DMPP (100 uM)- and McN-A-343 (100 uM)-induced CA secretions under existence of 17-╬▒-estradiol plus tamoxifen were also significantly recovered to 67.8┬▒3.1%(0ŌĆō4 min, p<0.01, n=8) and 59.7┬▒5.4%(4ŌĆō8 min, p<0.01, n=8) and 70.0┬▒4.8% (0.4 min, p<0.01, n=6) of their corresponding control responses, respectively, as compared to the secretory responses of 47.5┬▒2.5% (0ŌĆō4 min) and 21.7┬▒5.1% (4ŌĆō8 min), and 33.4┬▒3.5%(0ŌĆō4 min) of the control in the presence of 10 uM 17-╬▒-estradiol only (Fig. 11).
6. The Effect of 17-╬▒-estradiol Plus Metoclopramide on CA Release Evoked by ACh, Excess K+, DMPP and McN-A-343
It was tried to examine the relationship between dopaminergic and estrogenergic receptors in CA release. It has been reported that dopamine-induced inhibition of release of [3H] norepinephrine from the isolated perfused rabbit adrenal glands could be reversed completely by the dopamine D2 selective antagonist metoclopramide27,28).
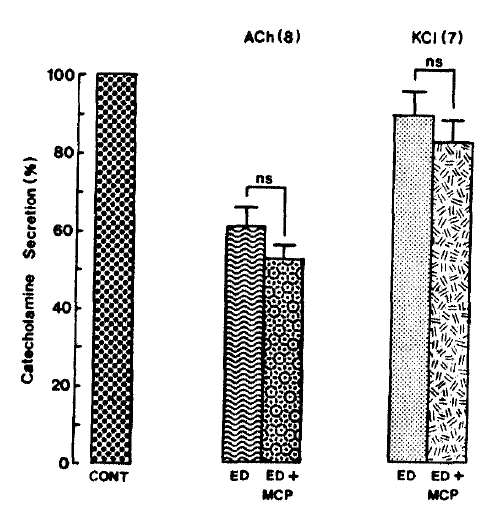
ACh (5.32mM)- and excess K+ (56 mM)-induced CA secretory responses after pre-loading with Krebs solution containing 10 uM 17-╬▒-estradiol along with 33 uM metroclopramide for 20 min amounted to 52.8┬▒3.2% and 82.3┬▒5.8% of each corresponding control (100%), respectively. They were not significant as compared to the secretory responses of 60.9┬▒4.6% and 89.1┬▒6.7% of the controls in the presence of 17-╬▒-estradiol only as shown in Fig. 13.
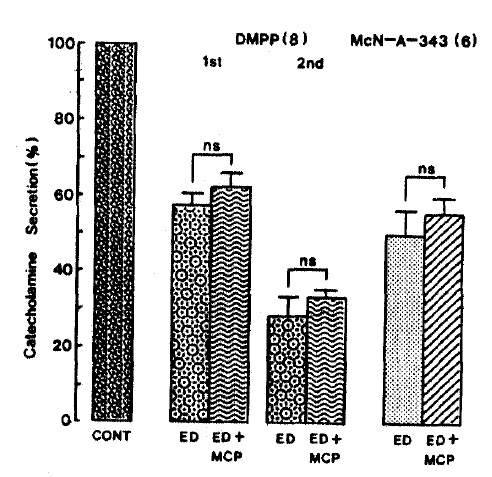
Furthermore, DMPP (100 uM)- and McN-A-343(100 uM)-stimulated CA secretory responses in the presence of 10 uM 17-╬▒-estradiol plus 33 uM metoclopramide were 62.4┬▒3.6% (0ŌĆō4 min, ns, n=8) and 33.2┬▒2.1% (4ŌĆō8 min, ns, n=8), and 55.9┬▒4.2% (0ŌĆō4 min. ns, n=6), respectively, as compared to their secretory responses of 57.3┬▒3.5% (0ŌĆō4 min) and 28.7┬▒5.1% (4ŌĆō8 min), and 50.2┬▒6.3% (0ŌĆō4 min) of the controls as depicted in Fig. 14.
DISCUSSION
In the present study, the experimental results suggest strongly that 17-╬▒-estradiol causes the inhibitory effect of CA secretion evoked by cholinergic (nicotinic or muscarinic) receptor stimulation and Bay-K-8644, but not that by membrane-depolarization, indicating that this effect may be mediated by inhibiting the influx of extracellular calcium into the rat adrenomedullary chromaffin cells through the activation of inhibitory estrogen receptors, and it also play a modulatory role in regulating CA secretion.
In support of this idea, Lopez and his collabolators13) have found that the steroid hormone 17-╬▒-estradiol blocks CA secretion from adrenal chromaffin cells in response to some secretagogues. Although this effect is especially clear upon nicotinic stimulation and, to a lesser extent with high K+ stimulation, it is dubious in the case of muscarinic-mediated secretion. However, it seems that there is some difference between results of previous reports and the present investigation. In the present experimental results, 17-╬▒-estradiol markedly inhibited CA secretory responses of DMPP and McN-A-343, but did not affect that of excess K+. The resson for this discrepancy is not explained, but the discrepancy may be due to the species differences in the adrenal medulla.
Moreover, Leret and his co-workers14) have also shown that, in the adrenal medulla, the CA levels measured in animals lesioned with 6-hydroxydopamine were significantly higher than those of the controls. Thus, it could be suggested that the CA of the amygdala plays an inhibitory role in animals with 6-hydroxydopamine lesions in the CNS region. The activity of the adrenal medulla can also be influenced by steroid hormones produced in the ovary29). These hormones not only modify the production of corticosteroids but also the activity of some medullary enzymes. Namely, it has been demonstrated that low levels of ovarian steroids reduce MAO and phenylethanolamine-N-methyltransferase (COMT) activities, with no changes of COMT activity14). This mechanism could be involved in the increase of CA in the adrenal medulla obtained after lesions in the amygdala. A similar effect has also been described in several brain areas30ŌĆō32). Thus, the adrenal medulla activity of COMT, the enzyme involved in the inactivation of CA by o-methylation, was significantly increased after the treatment with either estradiol or progesterone29).
Miguel and his co-workers12) have found that the CA release, measured after in vitro incubations of adrenomedullary tissue, is significantly reduced during the estrosus phase. Most likely, pre-ovulatory increase of estradiol, occurring during the early progestrous33), would be responsivle for this decreased CA release, since the pharmacological administration of estradiol to overiectomized rats produced a similar decrease in the CA release 24 hr after the steroid injection. They have also suggested that the effects of sex steroid on adrenal medulla would be produced by a direct action on chromaffin cells through a classic genomic mechanism34). In this respect, the existence of cytoplasmic steroid receptors has been observed in the whole adrenal35).
However, these results are not concordant with the present experimental data. Because, first, the inhibitory effect by 17-╬▒-estradiol of CA release is established within 20 min and, second, its effect is readily reversible upon washing out the hormone. Rather, its effect might be ascribed to the direct action to the membrane rapid actions observed in different neuronal systems. The rapid effect that, in the present investigation, 17-╬▒-estradiol inhibited CA secretion evoked by DMPP and McN-A-343 is not explained in the frame of the classical genomic mechanism of steroid hormone actions through activation of high-affinity cytosolic receptors and their further translocation to the cell nucleus to induce new protein synthesis. These effects are generally known to take hours or even days to appear36).
Rather, the catecholamine blocking effects are better explained in the newly emerging concept of cell membrane effects of steroids in neurons. In support of these ideas, there are several recent example reports of these nongenomic, rapid effects of steroids. By applying steroids directly to specific neurons, immediate changes in their firing frequency are observed10,11,37,38). The activity of hormone-sensitive striated muscles can be enhanced within minutes of steroid treatment39). As these rapid steroid effects are not prevented by protein synthesis inhibitors11,39), it seems that they result from a direct action of steroids on the plasma membrane. In the light of these facts, it is suggested that the target for 17-╬▒-estradiol is located on the plasma membrane of the rat adrenomedullary chromaffin cells. If this hormone were acting at some intracellular site on the secretory machinery, the responses to all secretagogues would be blocked the same extent.
Furthermore, in the present study, 17-╬▒-estradiol also inhibited the secretory effect of CA evoked by Bay-K-8644, which is known to be a Ca++uptake21) and CA release22). In terms of this view, the present finding that 17-╬▒-estradiol depressed CA secretory effects by Bay-K-8644, as well as cholinergic receptor-stimulation without effect on membrane-depolarization, strongly suggests that the inhibitory effect of 17-╬▒-estradiol is mediated through the blockade of Ca++ entry into the chromaffin cells.
In support of these results, there is now sizeable literature demonstrating a key role of Ca++ influx through voltage-sensitive Ca++ channels as a physiological pathway for activation of adrenal CA40ŌĆō45). Moreover, it is found that the activation of nicotinic receptors stimulates CA secretion by increasing Ca++ entry through receptor-linked and/or voltage-dependent Ca++ channels in both perfused rat adrenal glands46) and isolated bovine adrenal chromaffin cells47ŌĆō49), and that the muscarinic receptor activation causes an increase in adrenal CA secretion independent of extracellular Ca++ in various species50ŌĆō52) and in cytosolic free Ca++ in bovine isolated adrenal chromaffin cells without associated CA secretion45,53ŌĆō55).
However, recently, Lim and Hwang56) have found that both DMPP and McN-A-343 greatly cause CA secretion from the isolated perfused rat adrenal medulla by a calcium-dependent exocytotic mechanism. Considering in connection with these findings, the present results that 17-╬▒-estradiol inhibited CA releasing responses induced by DMPP and McN-A-343 as well as that by Bay-K-8644, but did not that by excess K+, indicate strongly that this inhibitory activity may be mediated by inhibiting Ca++ influx into chromaffin cells through receptor-linked and/or voltage-sensitive Ca++ channels. However, the fact that 17-╬▒-estradiol did not affect CA secretion evoked by high K+ could not be clearly explained, because DMPP and high K+ are known to increase Ca++ uptake into chromaffin cell through the recruitment of voltage-sensitive Ca++ channels28,57). In addition, Lopez and his co-workers13) have also observed that 17-╬▒-estradiol depresses the secretory responses of CA evoked by both DMPP and high K+ from the perfused bovine and cat adrenal medulla.
In terms of these reports, the present result that 17-╬▒-estradiol fails to inhibit CA release induced by excess K+ from the rat adrenal glands might explain its high potency in blocking CA secretion evoked by DMPP or McN-A-343 than by excess K+, indicating that there is species difference.
On the other hand, in this study, the fact that the inhibitory effect of 17-╬▒-estradiol to the secretory responses evoked by DMPP and McN-A-343 as well as by ACh was recovered to the considerable extent of the control in the presence of tamoxifen indicates strongly that this hormone causes depression in CA release evoked by cholinergic (nicotinic and/or muscarinic) receptor stimulation through the activation of inhibitory estrogen receptors on chromaffin cell membrane. Tamoxifen is a competitive inhibitor of estrogen binding at the estrogen receptor24ŌĆō25) and blocks estrogen action in breast cancer cells which contain receptors. Recently, Wiseman58) has described that tamoxifen protects membranes and lipoprotein particles against oxidative damage, and that this antioxidant action is likely to contribute to the observed cardio-protective action of tamoxifen and supports the use of this compound in treating and even preventing breast cancer. Membrane-mediated mechanisms of tamoxifen action, through a putative modulation of membrane fluidity are likely to play an important role in it anti-cancer action and its ability to reverse multi-drug resistance and could also lead to clinical uses as anti-candida and anti-viral agents.
Also, in the present investigation, 17-╬▓-estradiol, an isomer of estradiol, inhibited CA secretion evoked by cholinergic stimulation. In support of this fact, after 3 weeks of ovariectomy in female Clarias batrachus, CA levels were elevated significantly in the preparatory, pre-spawinig and spawning phases and were ulaltered in the post-spawning season23).
Administration of 17-╬▓-estradiol to this fish resulted in both seasonal and differential effects on the CA levels. Moreover, it has been reported that 17-╬▓-estradiol also produces brief hyperpolarization and increased potassium conductance in rat medial amygdala brain slices11).
Besides, the finding that 17-╬▒-estradiol-induced inhibitory effects were unchanged by co-administration with metoclopramide, which is known to block dopaminergic receptors, leading to hypertensive crisis59) and CA release by calcium-dependent exocytotic mechanism60), suggests that the inhibitory effects are not associated with dopaminergic receptors in the rat adrenal glands.
In conclusion, 17-╬▒-estradiol causes the inhibition of CA release evoked by cholinergic (nicotinic and/or muscarinic) stimulation as well as that by Bay-K-8644, but not that by membrane-depolarization from the isolated perfused rat adrenl glands, indicating strongly that this inhibitory effect may be mediated by inhibiting calcium influx into the chromaffin cells through the activation to the inhibitory estrogen receptors and it also plays a modulatory role in regulating CA secretion.


 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print





