


The Expression of β1 Integrin and Intercellular Adhesion Molecule-1 on Mesangial Cells and Its Modulation in Glomerulonephritis
- Chun Gyoo Ihm, M.D., Jae Phil Kim, M.D., Jae Kyung Park, M.D., Byung Jun Kim, M.D., Jae Hyung Ahn, M.D., Tae Won Lee, M.D., Moon Ho Yang, M.D., Myung Jae Kim, M.D.
- Abstract
-
- Objectives
- This study was performed to observe the effects of conditioned media (CM) of cultured mononuclear cells (MNC) on the expression of β1 integrin and intercellular adhesion molecule (ICAM)-1 on mesangial cells and the modulation by TGF-β. TNFα. or hydroxyl radical. Also these were examined in anti-Thy mesangial proliferative GN (MsPGN) or puromycin aminonucleoside nephrosis (PAN).
- Methods
- The expression of β1 integrin and ICAM-1 on mesangial cells was examined by ELISA and the modulation by TGF-β, TNF α, or hydroxyl radical was investigated by using neutralizing antibody and dimethylthiourea, [ · OH] scavenger.
- Results
- The CM of MNC from Control suppressed the β1 integrin expression and thymidine incorporation, and increased ICAM-1 expression on mesangial cells, compared to media alone, Especially the significant increase in ICAM-1 expression was reversed by anti-TNFα antibody. β1 integrin expression on mesangial cells was higher by the addition of CM of MNC in MsPGN than in Control, which was prevented by anti-TGF β antibody or dimethyl-thiourea. ICAM-1 expression was not different among groups. Thymidine incoporation by adding CM of MNC was lower in MsPGN and PAN, which was mitigated by anti-TGFβ and anti-TNFα. Thus the ratio of the expression of ICAM-1 to thymidine incorporation was higher in MsPGN and PAN.
- Conclusion
- The expression of β1 integrin and ICAM-1, and the thymidine incorporation on mesangial cells are directly regulated by MNC, maybe through factors such as TGFβ or TNFα. In MsPGN, the overexpression of β1 integrin induced by MNC through TGFβ or hydroxyl radical on day 3 may be related to the pathogenesis.
- INTRODUCTION
- INTRODUCTION
Although the pathogenesis has not been defined, a number of mediators have been thought to be candidates responsible for glomerulonephritis(GN)1). Mononuclear cells(MNC) have been reported to be identified in the glomeruli of patients with GN2,3). It’s suggested that MNC influx are associated with increased extracellular matrix(ECM) expression4). It has been reported that glomerular cells, especially mesangial cells, as well as MNC synthesize ECM proteins like collagen and fibronectin5), and release and re-spond to cytokine growth factors6–8). Transforming growth factor(TGF)β has marked effects on ECM accumulation by cultured mesangial cells9–12), while it inhibits proliferation of glomerular cells. The treatment with anti-TGF β antibody was reported to prevent proteoglycan production in experimental GN12). Tumor necrosis factor(TNF)α has the several effects on mesangial cells such as mesangiolysis, cell proliferation or procoagulant activity13,14). Serum TNFα and TNFα mRNA in kidney were found to be increased in GN15–18). Also, reactive oxygen species(ROS) released from leukocytes or glomerular cells, such as hydroxyl radical, were reported to be associated with proteolysis and prostaglandin synthesis, and to be related to some GN19).The expansion of mesangial ECM and cell proliferation appears to be important pathologic features in GN, and may contribute to the development of sclerosis20). It was reported that the antibodies to ECM proteins stain in the glomerular mesangium, and mRNAs for ECM protein are increased in several GN21–23). Recently, the adhesion molecules such as β1 integrin and intercellular adhesion molecules(ICAM)-1 have been detected in glomerulus24,25). They are suggested to play an important role as mediators of cell-matrix or cell-cell interactions in immune or inflammatory processes. The β1 integrin expression are known to be regulated by TGF β, and the ICAM-1 expression by TNFα or interleukin-1. Their expression was reported to be altered in some GN26–30).However, the direct relationships between MNC and the expression of adhesion molecules on mesangial cells have not been delineated yet.Therefore, this study was performed to observe the direct effects of conditioned media (CM) of cultured MNC on the expression of β1 integrin and ICAM-1 on mesangial cells, together with thymidine incorporation, and the modulation by TGF-β, TNFα, or hydroxyl radical on these was investigated. Also these were examined in anti-Thy mesangial proliferative GN(MsPGN) or puromycin aminonucleoside nephrosis(PAN).
- MATERIALS AND METHODS
- MATERIALS AND METHODS
- Induction of Experimental Models
- Induction of Experimental Models
Anti-Thy MsPGN was induced in Sprague-Dawley rats(5–6 wk old) by intravenous administration of 1 ml of anti-thymocyte antibody per 100g of body weight followed by 1 ml of normal rabbit serum12). Also PAN was induced by intravenous administration of 1.5mg of the aminonucleoside of puromycin per 100g of body weight31). The rats were killed on day 3 for histologic examination of kidney tissue and isolation of MNC for culture.- Rat Mononuclear Cell Culture
- Rat Mononuclear Cell Culture
Spleen cells were obtained from rats with Control, MsPGN or PAN and, after removal of erythrocytes, were cultured at 0.5×107 cells/ml of RPMI 1640 media supplemented with 10% heat-inactivated fetal calf serum in a humidified atmosphere of 5% CO2 in air at 37°C. After 48h, the CM were harvested, filtered, and applied to the cultured mesangial cells.- Rat Mesangial Cell Culture
- Rat Mesangial Cell Culture
Glomeruli were isolated using the graded sieving technique, and washed three times in PBS. For pure mesangial cell culture, collagenase treated glomeruli were plated on culture dishes in DMEM media containing 17% heat-inactivated fetal bovine serum, glutamine, penicillin, streptomycin, amphotericin B, and insulin. It was examined if these cells have prominent intracellular myosin fibrils, are negative with antibodies to rat la antigen and factor VII by immunofluorolescent staining as described, and are capable of growth in D-valine substituted medium and not sensitive to puromycin11,31). Near confluent cells in the third to fourth passage were used in these studies.- Assessment of Expression of β1 Integrin and ICAM-1 on Mesangial Cells
- Assessment of Expression of β1 Integrin and ICAM-1 on Mesangial Cells
1×104/well mesangial cells were plated into 96 well plates and cultured for 48h. After adding each components such as CM of MNC, antibodies against TGF β or TNFα(Genzyme, Cambridge, MA, USA) and a hydroxyl radical scavenger, dimethylthiourea, cells were cultured for 48 h and washed with DMEM twice. Each plates were fixed with 1% paraformaldehyde for 15 min at room temperature and washed. After blocking unbound region with 200μl of 2% bovine serum albumin, 100 μl of mouse ant-rat ICAM-1 antibody(Serotec, Oxford, England) and anti-rat β1 integrin antibody(Chemicon, Temecula, CA, USA) were added. Then alkaline phosphatase conjugated goat anti-mouse antibody(Gibco BRL) was added and cells were incubated for 1 h. After adding p-nitrophenyl phosphate, reaction was stopped with 3 N NaOH and the optical density(OD) was read at 405nm.- Thymidine Incorporation Assays
- Thymidine Incorporation Assays
Cells were plated at 1×104 cells per well in 96-well plates in DMEM medium. One μCi per well of [3H] thymidine(New England Nuclear, Boston, Massachusetts, USA) was added after the medium was replaced according to the experimental design as described above. After an 16h incubation, the contents of each well were counted in a liquid scintillation counter.- Statistics
- Statistics
Results are expressed as the mean ± standard deviation of the mean(SD). Comparison among groups were made by ANOVA using the Newman-Keuls multiple comparison test. Comparison between groups were made using Student’s paired t-test for normal distributed data. Significance was assigned at the p<0.05 levels.
- RESULTS
- RESULTS
- Experimental Glomerulonephritis
- Experimental Glomerulonephritis
On day 3 after the administration of anti-rat thymocyte serum, light microscopy showed mild hypercellularity in some tufts of glomeruli of Ms PGN rats. Also small electon-dense deposits were seen in the glomerular mesangia by electron microscopy. In PAN models the focal and segmental obliteration of foot processes was observed, and the swelling of the visceral epithelial cells with villous transformations became evident.- β1 Integrin Expression on Meangial Cells
- β1 Integrin Expression on Meangial Cells
The expression of β1 integrin was measured by ELISA on mesangial cells cultured for 48 h, after addition of CM of MNC harvested from rats with GN on day 3. The expression of β1 integrin on mesangial cells was decreased by 20%, compared to media alone, by the addition of CM of MNC from Control(Fig. 1). It was higher in MsPGN than in Control(n=6, 122 ± 12% vs. 102 ± 15%, p < 0.05)(Fig. 2). Further studies utilizing neutralizing antibody were perfomed to determine whether the above changes are modulated by TGF β. The increased expression was prevented by 25μg/ml anti-TGF β antibody(96 ± 11%)(Fig. 2). But, the treatment with anti-TGF β antibody also decreased the expression of β1 integrin in Control(71 ± 10%). Marginal increments in the expression of β1 integrin was noticed in PAN(113 ± 10%)(p<0.05).- ICAM-1 Expression on Mesangial Cells
- ICAM-1 Expression on Mesangial Cells
The expression of ICAM-1 was increased by 44% by the addition of CM of MNC(Fig. 1). While there were no differences among groups (Table 1), the ratio of the expression of ICAM-1 to thymidine incorporation was higher in MsPGN and PAN than in Control. Further studies utilizing neutralizing antibody showed that the increased expression was completely prevented by 50μl/ml anti-TNFα antibody(Table 1), and TNFα stimulated the expression of ICAM-1 by 33%.- Thymidine Incorporation Assay on Mesangial Cells
- Thymidine Incorporation Assay on Mesangial Cells
[3H] thymidine incorporation was examined to observe the cell growth in relation to β1 integrin expression or cytokine growth factors. It was greatly suppressed by adding CM of MNC from Control(Fig. 1). Also it was lower in MsPGN(43±5×102cpm), and PAN(37±7×102cpm) than in Control(122 ± 20 × 102cpm), which was mitigated by anti-TGFβ and anti-TNFα(Table 2). Especially the thymidine incorporation in MsPGN was reversed to normal by anti-TNFα(101±9×102cpm).- Hydroxyl Radical Scavenger Treatment
- Hydroxyl Radical Scavenger Treatment
To investigate the relationship between hydroxyl radical and the above changes, a hydroxyl radical scavenger, 10mmol/l dimethylthiourea, was added. Then, the increased expression of β1 integrin induced with CM of MNC from MsPGN was prevented(98 ± 8%). There were no changes in ICAM-1 expression after dimethylthiourea treatment. The decrease in thymidine incorporation induced with the CM of MNC from MsPGN was mitigated with dimethylthiourea(43 ± 5 × 102cpm vs. 64±4×102cpm)(p<0.05).
- DISCUSSION
- DISCUSSION
These experiments were perfomed to test the hypothesis that 1) the CM of MNC affects the expression of β1 integrin or ICAM-1 on mesangial cells, which may mediate the ECM accumulation and cell proliferation in glomeruli leading to glomerulosclerosis in glomerulonephritis; and 2) some cytokine growth factors, or hydroxyl radical released by MNC have the central role in a series of these changes.According to our data, the CM of MNC from Control suppressed the β1 integrin expression and thymidine incorporation. Especially it increased ICAM-1 expression on mesangial cells by 44%. This study showed that MNC may have the important factors regulating mesangial cell phenotype. The above changes were in part ascribed to the effects of TGFβ or TNFα. The increase in ICAM-1 expression was prevented by anti-TNFα.In anti-Thy MsPGN, mesangiolysis occurs by the formation of in situ immune complex during the first 1 or 2 days, and later is followed by cell proliferation32,33). PAN is the model of nephrotic syndrome caused by epithelial cell injury18,34). In this study, the expression of β1 integrin on mesangial cells was increased by the CM of MNC from MsPGN group on day 3. The β1 subunit associates with multiple α’s, comprising at least nine integrin. The β1 integrin binds to the ligands such as collagen, laminin, fibronectin or Arg-Gly-Asp sequence, and can promote cell adhesion to ECM known to accumulate in GN. There is evidence that the β1 integrin affects matrix metabolism24). The binding of mesangial cells to collagen IV, via the β1 integrin, stimulates the original construction of new matrix starting at the cell-matrix contact site. Also it’s suggested that MNC may have the central role in the regulation of β1 integrin expression in anti-Thy MsPGN, consistent with the fact that this GN is monocyte dependent32). TGF β is known to stimulate the production of β1 integrin, together with ECM protein, contributing to matrix expansion or glomerulosclerosis10,24). The treatment with anti-TGF β antibody prevented the overexpression of β1 integrin induced with the CM of MNC from MsPGN in this study. These in vitro data are further consistent with the recent report that β1 integrin expression is coordinated with TGF β-induced ECM protein synthesis in glomeruli of rats with anti-Thy MsPGN30). Moreover, our experimental methods may be beneficial to test the direct relationships between β1 integrin and MNC in GN. However, the increase in β1 integrin expression was small and anti-TGF β antibody also suppressed the β1 integrin expression induced by the CM of MNC from control. This cannot rule out that the other factors can also be candidates for these changes. Our preliminary data showed interleukin-1 β increased the expression of β1 integrin by 24%, which is also supported by other work35), but in conflict with a recent report30).The expression of ICAM-1 on mesangial cells was increased by 44% by the addition of CM of MNC from Control, and was completely prevented by anti-TNFα antibody. Also our experiments showed that TNFα stimulated the expression of ICAM-1, consistent with other studies25,36). Altogether this supports the role of TNFα produced by MNC on ICAM-1 expression. However, in these experiments ICAM-1 expression was not different among groups. This might be explained by the decreased cell viability due to mesangiolysis in MsPGN or the presence of certain factors other than TNFα. The ratio of the expression of ICAM-1 to thymidine incorporation was higher in GN groups than in control.While the thymidine incorporation was suppressed by the addition of CM of MNC from Control, it was lower in MsPGN and PAN than in Control. The decrease in thymidine incorporation was mitigated by anti-TGF β or anti-TNFα. The findings suggest that the decreased cellularity by CM of MNC may be, in part, due to anti-proliferative action of TGF β or cytolytic effects of TNFα11,13). Especially the thymidine incorporation in MsPGN was reversed to normal by anti-TNFα. Also one may speculate that there may be the close relationships between adhesion molecules, some cytokine growth factors, and cellularity on mesangial cells in GN37). In our experiments the decreased cellularity was seen concurrent with the increase in β1 integrin in MsPGN.A hydroxyl radical scavenger treatment prevented the overexpression of β1 integrin induced by CM of MNC from MsPGN group, like anti-TGF β. Because cytokines like interleukin-1 and TNFα stimulate ROS generation in mesangial cells38), the relationships between hydroxyl radical and the expression of β1 integrin, through cytokines, can be speculated.In conclusion, the expression of β1 integrin and ICAM-1, and the thymidine incorporation on mesangial cells are directly regulated by MNC, maybe through factors such as TGF β or TNFα. In MsPGN, the overexpression of β1 integrin induced by MNC through TGF β or hydroxyl radical on day 3 may be related to the pathogenesis. Further studies are necessary to investigate the expression of β1 integrin and ICAM-1 in GN at the late time points, and also the modulation by other cytokine growth factors or systems such as protein kinase C or eicosanoid.
- Acknowledgments
- Acknowledgments
This investigation was supported by grants from the KoWhang foundation of KyungHee University.
Fig. 1.
Effects of the conditioned media of mononuclear cells on the expression of β1 integrin and ICAM-1, and[3H] thymidine incorporation. The conditioned media of mononuclear cells from Control(■) suppressed the β1 integrin expression and thymidine incorporation, and increased ICAM-1 expression on mesangial cells, compared to media alone(□). a : p<0.05 vs. media alone.
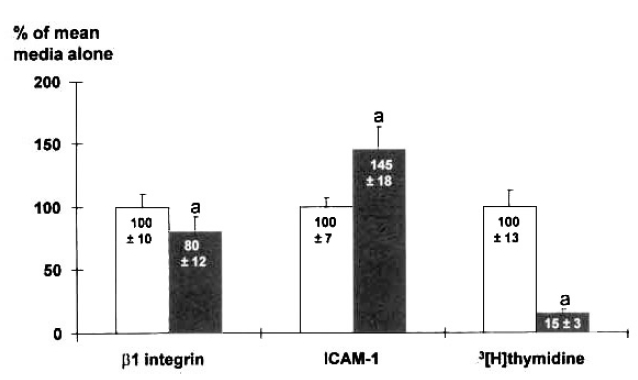
Fig. 2.
The expression of β1 integrin on mesangial cells after the addition of conditioned media of mononuclear cells harvested from rats with glomerulonephritis. The expression in MsPGN was higher than in Control. The overexpression was prevented by the tratment with 25μg/ml anti-TGF β antibody(■), which also decreased the β1 integrin expression in Control. a : p < 0.05 vs. Control, b : p < 0.05 vs. without anti-TGF β antibody(□).
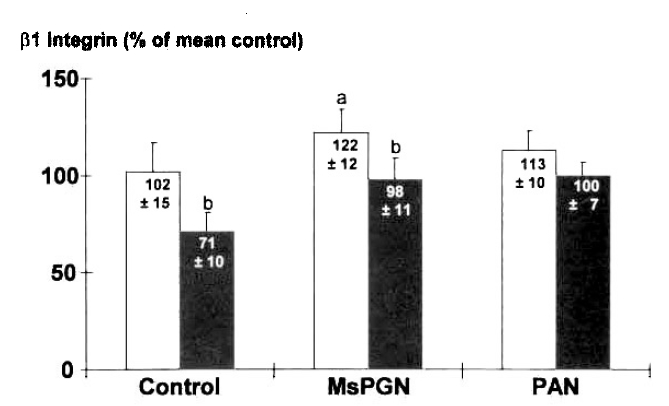
Table 1.
Effect of the Conditioned Media of Mononuclear Cells on the Expression of ICAM-1 on Mesangial Cells
| Mononuclear cellconditioned media |
ICAM-1(% of mean control value)
|
|
|---|---|---|
| without anti-TNFα with anti-TNFα | ||
| Control | 100 ± 12 | 72 ± 11a |
| MsPGN | 102 ± 7 | 69 ± 14a |
| PAN | 94 ± 8 | 75 ± 11a |
Table 2.
Effect of the Conditioned Media of Mononuclear Cells on[3H] Thymidine Incorporation on Mesangial Cells
| Mononuclear cell conditioned media |
[3H] thymidine incorporation(102cpm)
|
||
|---|---|---|---|
| without | with anti-TNFα | with anti-TGFβ | |
| Control | 122 ± 20 | 170 ± 6b | 141 ± 15 |
| MsPGN | 43 ± 5a | 101 ±9b | 64 ± 8b |
| PAN | 37 ± 7a | 56 + 6b | 49 ± 5b |
- References
- References
REFERENCES
1. Couser WG. Mechanisms of glomerular injury in immune complex disease. Kidney Int 28:569–5831985.
[Article] [PubMed]2. Arima S, Nakayama M, Naito M, Sato T, Takayashi K. Significance of mononuclear phagocytes in IgA nephropathy. Kidney Int 39:684–6921991.
[Article] [PubMed]4. Floege J, Alpers CE, Burns MW, Pritzl P, ordon K, Couser WG, Johnson RJ. Glomerular cells, extracellular matrix accumulation, and the development of glomerulosclerosis in the remnant kidney model. Lab Invest 66:485–4971992.
[PubMed]5. Ishimura E, Sterzel RB, Budde K, Kashgarian M. Formation of extracellular matrix by cultured rat mesangial cells. Am J Pathol 134:843–8551989.
[PubMed] [PMC]6. Shultz PJ, Raji L. The glomerular mesangium. Am J Kidney Dis(Suppl) 17:8–141991.7. Wardle EN. Cytokine growth factors and glomerulonephritis. Nephron 57:257–2611991.
[Article] [PubMed]8. Kujubu DA, Fine LG. Physiology and ceil biology update: Polypeptide growth factors and their relation to renal disease. Am J Kidney Dis 14:61–731989.
[Article]9. MacKay K, Striker LJ, Stauffer JW, Agodoa LY, Striker GE. Transforming growth factor-β. J Clin Invest 83:1160–11671989.
[Article] [PubMed] [PMC]11. Border WA, Okuda S, Languino LR, Ruoslahti E. Transforming growth factor-β regulates production of proteoglycans by mesangial cells. Kidney Int 37:689–6951990.
[Article] [PubMed]12. Okuda S, Languino LR, Ruoslahti E, Border WA. Elevated expression of transforming growth factor-β and proteoglycan production in experimental glomerulonephritis. J Clin Invest 86:453–4621990.
[Article] [PubMed] [PMC]13. Jaattela M. Biologic activities and mechanisms of tumor necrosis factor/cachectin. Lab Invest 64:724–7421991.
[PubMed]14. Meulders Q, He CJ, Adida C, Peraldi MN, schleuning WD, Sraer JD, Rondeau E. Tumor necrosis factor α increases antifibrinolytic activity of cultured human mesangial cells. Kidney Int 42:327–3341992.
[Article] [PubMed]15. Suranyi MG, Guasch A, Hall BM. Elevated levels of tumor necrosis factor-α in the nephrotic syndrome in humans. Am J Kidney Dis 21:251–2591993.
[Article] [PubMed]16. Noronha IL, Kruger C, Andrassy K, Ritz E, Waldherr R. In situ production of TNF-α, IL-β and IL-2R in ANCA-positive glomerulon-ephritis. Kidney Int 43:482–6921993.
[Article]17. Pan CG, Bresnahan BA, Albrightson C, Lianos EA. Mesangial cell immune injury: Synthesis, origin, and role of tumor necrosis factor(TNF). J Am Soc Nephrol 4:627. 1993;(Abstract).18. Diamond JR, Pesek I. Glomerular tumor necrosis factor and interleukin 1 during acute aminonucleoside nephrosis. Lab Invest 64:21–281991.
[PubMed]19. Diamond JR. The role of reactive oxygen species in animal models of glomerular disease. Am J Kidney Dis 19:292–3001992.
[Article] [PubMed]20. Klahr S, Schreiner G, Ichikawa I. The progression of renal disease. N Engl J Med 318:1657–16661988.
[Article] [PubMed]21. Alder S, Striker J, Striker GE, Perkinson DT, Hibbert J, Couser WG. Studies of progressive glomerular sclerosis in the rat. Am J Pathol 123:553–5621988.22. Striker LM, Killen PD, Chi E, Striker GE. The composition of glomerulosclerosis. I. studies in focal sclerosis, crescentic glomerulonephritis, and membranoproliferative glomerulonephritis. Lab Invest 51:181–1921984.
[PubMed]23. Flooege J, Johnson RJ, Gordon K, Iida H, Pritzl P, Yoshimura A, Campbell , Alpers CE, Couser WG. Increased synthesis of extracellular matrix in mesangial proliferative nephritis. Kidney Int 40:477–4881991.
[Article] [PubMed]24. Cosio FG. Cell-matrix adhesion receptors: Relevance to glomerular pathology. Am J Kidney Dis 20:294–3051992.
[Article] [PubMed]25. Wuthrich RP. Intercellular adhesion molecules and vascular cell adhesion molecule-1 and the kidney. J Am Soc Neph 3:1201–12111992.
[Article]26. Wuthrich RP. Vascular cell adhesion molecule-1 (VCAM-1) expression in murine lupus nephritis. Kidney Int 42:903–9141992.
[Article] [PubMed]27. Canton AD, Fuiano G, Sepe V, Caglioti A, Ferrone S. Mesangial expression of intercellular adhesion molecule-1 in primary glomer-ulosclerosis. Kidney Int 41:951–9551992.
[Article] [PubMed]28. Lhotta K, Neumayer HP, Joannidis M, Geissler D, Konig P. Renal expression of intercellular adhesion molecule-1 in different forms of glomerulonephritis. Clinical Science 81:477–4811991.
[Article] [PubMed]29. Muligan MS, Johnson KJ, Todd RF, Issekutz TB, Miyasaka M, Tamatan T, Smith CW, Anderson DC, Ward PA. Requirements for leukocyte adhesion molecules in nephrotoxic nephritis. J Clin Invest 91:577–5871993.
[Article] [PubMed] [PMC]30. Kagami S, Border WA, Ruoslahti E, noble NA. Coordinated expression of β1 integrins and TGF-β 1-induced matrix proteins in glomer-ulonephritis. Lab Invest 69:68–761993.
[PubMed]32. Stahl RAK, Thaiss F, Kahf S, Schoeppe W, Helmchen UM. Immune-mediated mesangial cell injury-Biosynthesis and function of prostanoids. Kidney Int 38:273–2811990.
[Article] [PubMed]33. Yamamoto T, Wilson CB. Complement dependence of antibody-induced mesangial cell injury in the rat. J Immunol 138:3758–37651987.
[Article] [PubMed]34. Groggel GC, Hughes ML. Production of an inhibitor of rat mesangial cell growth by the glomerulus and its alteration in puromycin nephrosis. J Am Soc Nephrol 4:155–1611993.
[Article] [PubMed]35. Dehar S. Regulation of expression of the cell adhesion receptors, integrins, by recombinant human interleukin-1β in human osteosarcoma cells: Inhibition of cell proliferation and stimulation of alkaline phosphatase activity. J Cell Physiol 138:291–2991989.
[Article] [PubMed]36. Rothlein R, Czajkowski M, O’Neill MM, Marlin SD, Mainolfi E, Merluzzi VJ. Induction of intercellular adhesion molencule-1 on primary and continuous cell lines by pro-inflammatory cytokines. J Immunol 141:1665–16691988.
[PubMed]