New targets for type 2-low asthma
Article information
Abstract
Asthma is characterized by airway obstruction and inflammation, and presents significant diagnostic and treatment challenges. The concept of endotypes has improved understanding of the mechanisms of asthma and has stimulated the development of effective treatment strategies. Sputum profiles may be used to classify asthma into two major inflammatory types: type 2-high (T2H) and type 2-low (T2L) asthma. T2H, characterized by elevated type 2 inflammation, has been extensively studied and several effective biologic treatments have been developed. However, managing T2L is more difficult due to the lack of reliable biomarkers for accurate diagnosis and classification. Additionally, conventional anti-inflammatory therapy does not completely control the symptoms of T2L; therefore, further research is needed to identify effective biologic treatments. This review provides new insights into the clinical characteristics and underlying mechanisms of severe T2L and investigates potential therapeutic approaches to control the disease.
INTRODUCTION
Asthma is a respiratory disease characterized by airway obstruction and immune cell infiltration, as well as complex diagnostic and therapeutic requirements [1,2]. The concept of endotypes has improved our understanding of the underlying functional and pathophysiological processes involved in the condition, and has facilitated the identification of distinct subtypes within patient populations.
Among the various endotypes, two major categories have been identified: type 2-high (T2H) and type 2-low (T2L) [3–5]. The former, which is commonly associated with eosinophil activation and characterized by elevated levels of type 2 inflammation, is more extensively studied and well-defined. In general, clinicians can control and manage T2H by using steroid medications and targeted biologic interventions. However, treating T2L is more challenging due to heterogenous phenotypes and endotypes and the lack of reliable biomarkers [6–8].
Recent studies have investigated the inflammatory cytokines and chemokines associated with neutrophil activation in severe asthma (SA), including interleukin (IL)-1β, IL-6, IL-8, IL-17, IL-23, and tumor necrosis factor (TNF)-α. Novel interventions have been developed to modulate neutrophil infiltration and activation and to restore sensitivity to corticosteroids. Although these approaches may improve lung function and symptom control in patients with T2L, they may be difficult to implement in widespread clinical practice [9]. Here, we review the molecular mechanisms associated with severe T2L, identify potential biomarkers, and discuss the current and future treatment options for patients with this condition.
CLINICAL CHARACTERISTICS OF T2L
Various methods may be used to differentiate between T2L and T2H endotypes, and each method has its strengths and weaknesses. Previously, T2L was identified by the absence of specific inflammatory and immunological biomarkers associated with T2H, such as high levels of fractional exhaled nitric oxide, increased levels of eosinophils in the blood or sputum, and elevated levels of immunoglobulin E or periostin in serum. This simple noninvasive method enabled researchers to identify some of the unique characteristics of T2L, including its higher prevalence among older individuals with late-onset asthma, its resistance to standard medications, and its propensity for asthma exacerbations and increased severity [9–11]. However, various cutoff values were used for the biomarkers used to classify T2H, resulting in a wide range of interpretations in identifying T2L. Additionally, the presence of both eosinophils and neutrophils in patients with T2H is closely linked to an increased risk for asthma exacerbations and a poor response to steroid treatment. This suggests that the current classification system may be of limited value in identifying suitable treatments and predictive biomarkers [7,9,12]. Therefore, alternative methods for categorizing asthma phenotypes have been considered, such as analyzing patterns of cellular inflammation and sequestration in the lungs. Consequently, T2L may be classified as neutrophilic asthma (NA; with sputum neutrophilia), mixed granulocytic asthma (MGA; with both eosinophilia and neutrophilia in the sputum), or paucigranulocytic asthma (PGA; with no evidence of sputum eosinophilia or neutrophilia) [7,8]. The prevalence, diagnosis, features, and biomarkers associated with each of these phenotypes are discussed in the following sections.
Neutrophils constitute approximately 50–70% of circulating leukocytes in humans, and they play a critical role in defense against microbial invaders. However, patients with NA or MGA may exhibit both an increase in the level of neutrophils and impaired neutrophil function. Importantly, neutrophil levels in the blood may not accurately reflect neutrophil levels in the sputum [9,13,14]. Therefore, both inflammatory patterns in the sputum and biopsy samples must be evaluated. Various cutoff values for neutrophil counts in sputum samples, ranging from 40 to 76%, have been suggested to define NA, considering factors such as age, sputum induction, collection techniques, demographics, and previous exposure to microbiota and air pollutants [3,15–19]. As a result, patients with NA and those with MGA may include 4.3–5.4% and 5.2–6.7% of all patients with asthma, respectively [8,20]. Notably, Shi et al. [8] identified patients with NA or MGA using a cutoff value of 61% and discovered that these patients exhibited low levels of nitric oxide, reduced blood eosinophil and basophil counts, impaired lung function, frequent neutrophilia, and an increased requirement for high-dose inhaled corticosteroids compared to those with eosinophilic asthma. Although the appropriate cutoff values for predicting active neutrophil involvement in the pathogenesis of asthma remain unclear, the focus on neutrophils is important for both clinical practice and research.
The gold standard for evaluating airway inflammation is bronchial biopsy, which provides a comprehensive assessment when combined with sputum inflammatory cell profiles. A previous study employed these methods to examine the clinical, functional, and biological aspects of NA in 70 patients with mild to SA [21]. The authors established a cutoff value of 47.17 neutrophils per square millimeter in the bronchial lamina propria to define NA among these patients [22]. When NA was evaluated by induced sputum analyses, distinctive demographic and clinical characteristics were consistently observed. These characteristics included increased risks of asthma severity and exacerbations, a reduced response to bronchodilators (as indicated by decreased forced expiratory volume [FEV]1% and functional markers indicating airway closure), air trapping (as measured according to forced vital capacity [FVC]%), and an increased functional residual capacity. Moreover, these patients required higher doses of inhaled and oral corticosteroids to control their symptoms.
The methods for differentiating between T2H and T2L have considerable limitations, including the absence of reliable cutoff points, the need for specialized equipment, and invasiveness [21]. Therefore, these approaches are not suitable for all patients, and serum biomarkers may be used instead. One such biomarker that may be used to identify patients with NA is chitinase-like protein, also known as YKL-40 or human cartilage glycoprotein 39 [22]. Studies have revealed increased levels of this molecule in patients with asthma, and these levels positively correlate with YKL-40 levels in the lung, reduced FEV1%, and increased risk for SA. Furthermore, serum levels of YKL-40 are significantly correlated with NA and markers of neutrophil activation, including myeloperoxidase, IL-8, IL-6, and soluble IL-6 receptor [22,23]. In addition, we previously identified two biomarkers, S100 calcium-binding protein A9 and amyloid A1 in serum, that may be used to identify patients with NA [3,4], although further studies are needed to validate those findings.
Approximately 17–48% of asthma cases exhibit the PGA phenotype, which is characterized by normal levels of eosinophils in peripheral blood and eosinophils and neutrophils in sputum, together with the lowest nitric oxide levels among the various phenotypes [8,9,15,20]. PGA is considered the mildest form of asthma, showing favorable responses to appropriate anti-inflammatory medications [8,15,20]. However, among patients with PGA, 21.7% still experience severe refractory asthma and 14.8% record Asthma Control Test scores < 19, suggesting suboptimal disease management [24]. Furthermore, the lack of a consensus on well-defined characteristics for PGA has made it difficult to understand the underlying mechanisms and to develop targeted treatment strategies for this phenotype.
Impaired lung function in patients with PGA is associated with oxidant/antioxidant imbalance in the lungs. Notably, changes in the levels of glutaredoxin-1 and protein-glutathione mixed disulfides in sputum can distinguish patients with PGA from those with other asthma phenotypes [25]. Furthermore, distinct asthma phenotypes may now be identified using transcriptomic analyses of sputum samples, which can identify characteristic gene expression patterns. This approach has identified three groups that are similar to those previously identified by analyzing induced sputum: TAR1 (eosinophilic asthma), TAR2 (NA), and TAR3 (PGA) [26,27]. The application of sputum transcriptomics has the potential to enhance our ability to detect and differentiate among various asthma phenotypes including PGA. These findings have improved our understanding of the relevant pathophysiological mechanisms and have provided a basis for PGA treatment strategies.
NA AND MGA: MECHANISMS AND THERAPEUTIC TARGETS
Steroids may have anti-apoptotic effects on neutrophils, resulting in patients with NA or MGA exhibiting poorer responses to steroid treatment. Therefore, reliable biomarkers that can guide personalized treatment strategies are essential for managing NA and MGA. Although both human and animal studies have provided valuable insights into immune cell activation and cytokine production in neutrophil recruitment and activation, few biomarkers have been identified. The relevant immune cell populations, signaling pathways, and symptom control strategies are summarized in Table 1 [28–46] and Figure 1.

Potential therapeutic strategies for T2L
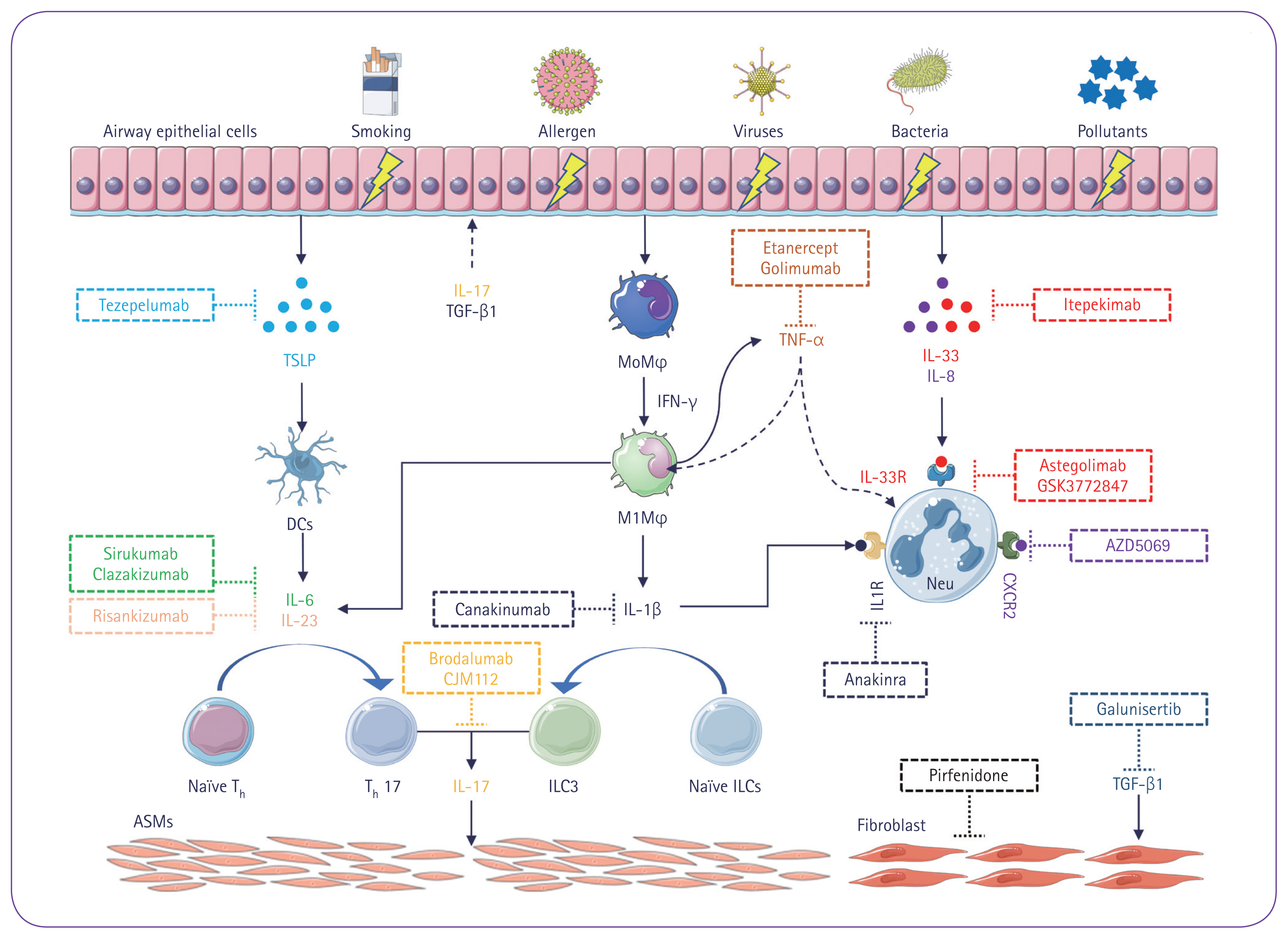
Mechanisms of airway inflammation and remodeling in T2L. Following exposure to various environmental triggers, such as viruses, bacteria, smoking, pollutants, and allergens, airway epithelial cells are activated and release the alarmins TSLP and IL-33, as well as IL-8. These cytokines mediate the migration and activation of neutrophils. In addition, increased levels of IFN-γ lead to macrophage activation and polarization, which stimulate neutrophilic airway inflammation. Activated macrophages and dendritic cells release the cytokines IL-6, IL-1β, and IL-23, which mediate differentiation of Th cells into Th17 and ILCs into ILC3. This results in the overproduction of IL-17, which in turn promotes airway remodeling by stimulating airway smooth muscle cell hyperplasia, tissue hypertrophy, and the release of IL-8 and TNF-α, thereby activating neutrophils. Furthermore, overproduction of TGF-β1 induces fibroblast proliferation, contributing to airway remodeling in T2L. Images were created using Servier Medical Art software (https://smart.servier.com/), licensed under a Creative Commons Attribution 3.0 Unported License (https://creativecommons.org/licenses/by/3.0/). ASM, airway smooth muscle; CXCR2, C-X-C motif chemokine receptor 2; DCs, dendritic cells; IL, interleukin; ILCs, innate lymphoid cells; IFN, interferon; IL-33R, IL-33 receptor; Mϕ, macrophages; Neu, neutrophils; T2L, type 2-low; TGF, transforming growth factor; Th, T helper; TNF, tumor necrosis factor; TSLP, thymic stromal lymphopoietin.
Therapies targeting the epithelium
Airway epithelial cells (AECs) play crucial roles in communicating between the intracellular and extracellular environments via cytokine signaling. When AECs encounter cellular injury, infection, irritants, or pollution, they produce alarmins that include IL-33 and thymic stromal lymphopoietin (TSLP). Moreover, AECs release C-X-C motif chemokine ligand 8 (CXCL8, also called IL-8), a cytokine that recruits and activates neutrophils, particularly in patients with SA.
Alarmins, such as IL-33 and TSLP, initiate downstream signaling pathways upon release. Long variants of TSLP selectively bind to their cognate receptors and the IL-7Rα subunit [28,29,47]. Activation of these receptors initiates various signaling pathways, resulting in immune cell activation [48]. TSLP can induce dendritic cell activation to release the cytokines IL-6 and IL-23, which are critical for the differentiation of immature CD4+ T cells into T helper (Th)17 lymphocytes. Neutrophil activation involves a process called “NETosis,” in which neutrophils undergo morphological changes in response to bacterial, fungal, viral, or inflammatory triggers [28,29,47,49]. This process leads to the release of extracellular chromatin fibers that contain granular proteins and enzymes, forming neutrophil extracellular traps (NETs). These NETs form a reticulate structure that traps and neutralizes pathogens. Furthermore, they have immunomodulatory effects and can activate monocytes, macrophages, and dendritic cells, thereby enhancing immune surveillance and antigen presentation. However, the unregulated release or persistence of NETs can exacerbate autoimmune diseases, thrombosis, tissue damage, and conditions such as NA or MGA [50]. Importantly, the presence of NETs may damage AECs and activate eosinophils. IL-33 can trigger neutrophil activation and release NETs by binding to IL-1 receptor-like 1/suppression of tumorigenicity (ST)2 and its coreceptor, IL-1 receptor accessory protein [47,79,51]. Severe uncontrolled asthma may be treated by neutralizing antibodies that target these alarmins but further studies are needed to determine how this affects neutrophilic airway inflammation [28,29].
Increased expression of the IL-33 receptor and IL-33-activated gene signatures have been observed in patients with severe NA and/or MGA. In addition, clinical trials targeting the IL-33 receptor ST2 have produced positive results. For example, treatment with astegolimab, which specifically inhibits the IL-33 receptor, resulted in decreased airway neutrophilic inflammation in patients with SA including those with T2L [30]. Similar outcomes were achieved in a separate study that used the inhibitor GSK3772847 [31]. These findings highlight the potential for targeting the IL-33/ST2 pathway as a therapeutic strategy to reduce neutrophilic airway inflammation in patients with severe uncontrolled asthma.
IL-8 plays an important role in stimulating neutrophil recruitment to airways and inducing the production of NETs in patients with SA [49]. Furthermore, the level of IL-8-induced NETs is negatively correlated with the FEV1/FVC ratio, indicating a potential link between NET production and airflow limitation. In addition, IL-8 recruits circulating monocytes to the airways; these can differentiate into various types of macrophage, thereby increasing neutrophilic airway inflammation [9]. Because IL-8 is clearly involved in the pathogenesis of T2L, targeting IL-8 is a potential therapeutic approach. Although there are no specific antibodies that neutralize IL-8, studies have focused on blocking its receptor: C-X-C motif chemokine receptor (CXCR)2. Two randomized controlled clinical trials evaluated the therapeutic effects of the human CXCR2 chemokine receptor inhibitors AZD6059 and SCH 527123. These studies demonstrated that treatment with these inhibitors reduced sputum neutrophil counts in patients with NA but did not result in significant improvements in clinical outcomes [32,33]. Although further research is needed to understand the role of IL-8 in neutrophilic airway inflammation and to develop specific IL-8 neutralizing antibodies, blocking the IL-8 signaling pathway via CXCR2 inhibition may reduce neutrophilic airway inflammation in patients with SA.
Therapies targeting leukocytes
The differentiation of naïve CD4+ T cells into distinct subsets, such as Th1, Th2, and Th17 cells, is crucial for driving disease mechanisms in different asthma endotypes. Patients with T2H exhibit increased levels of type 2 cytokines (such as IL-4, IL-5, and IL-13 released from Th2 cells), which are positively correlated with eosinophil levels in sputum and blood [52]. Conversely, the activation of neutrophils and macrophages in patients with T2L may be due to the increased production of interferon (IFN)-γ from Th1 cells and of cytokines (such as IL-17A, IL-17E, IL-17F, and IL-22) from Th17 cells. These factors collectively contribute to the recruitment and activation of neutrophils, a decline in lung function, and a decline in steroid sensitivity [52]. In addition to CD4+ T cells, innate lymphoid cells (ILCs), including ILC1, ILC2, and ILC3, also play important roles in stimulating asthma pathogenesis. ILCs are tissue-resident cells and do not possess lineage markers on their cell surfaces, unlike T cells [53]. However, ILCs can produce similar levels of cytokines. Understanding the cellular mechanisms responsible for the differentiation and activation of these Th and ILC cell subsets is crucial for developing targeted therapies for specific asthma endotypes.
Type 1 (IFN-α and IFN-β), type 2 (IFN-γ), and type 3 (IFN-λ) IFNs can combat viral and bacterial infections due to their antiviral, antiproliferative, and immunomodulatory properties [54,55]. IFN-α, IFN-β, and IFN-λ are predominantly produced in epithelial and hematological cells, whereas IFN-γ is primarily released by immune cells, including Th1, ILC1, CD8+ T, and natural killer T cells, as well as by macrophages [55]. Impaired innate immunity may be associated with deficiencies in genes involved in IFN-β and IFN-λ signaling, leading to acute asthma attacks. No clinical trials have investigated whether IFN-λ administration is beneficial for patients with SA, but the results of IFN-β administration have been somewhat disappointing [34,35]. Interestingly, there is a greater proportion of IFN-γ+ CD4+ T cells present in the airways of patients with SA than in patients with mild to moderate asthma [56]. Moreover, patients with poorly controlled NA exhibit elevated levels of IFN-γ produced by natural killer cells, indicating the involvement of a type 1 immune response in neutrophilic airway inflammation [57]. IFN-γ affects neutrophilic airways via the following mechanisms: increasing the production of CXCL10, which recruits Th1-polarized CD4+ T cells and neutrophils into the airways; reducing the production of secretory leukocyte protease inhibitor by AECs, leading to increased airway hyperresponsiveness (AHR), mast cell tryptase production, and steroid resistance; and activating phagocytes, which recruit and activate Th1 cells and ILC1 [57–59]. However, despite the importance of IFN-γ in neutrophilic airway inflammation, few therapeutic targets have been identified.
IL-17 is mainly released by Th17 and ILC3 cells, and this is regulated by the nuclear hormone receptor retinoic acid-related orphan receptor (ROR)-γt in mice and ROR-c in humans as well as by signal transducer and activator of transcription 3 [53]. Increased levels of IL-17 have been observed in patients with severe NA and are associated with bacterial infection, neutrophilia, smoking, frequent exacerbations, steroid resistance, and oral corticosteroid dependence [21,53,60,61]. IL-17 contributes to airway remodeling by promoting goblet cell hyperplasia, airway smooth muscle cell hyperplasia, and tissue hypertrophy. It also stimulates structural cells to release CXCL8/IL-8, granulocyte macrophage colony-stimulating factor, and TNF-α, which in turn activate neutrophils [53]. Moreover, cells producing IL-17 can be activated by cytoplasts generated during NET formation and by chemokine ligand (CCL)2 and CCL20 released during the interaction between IFN-γ and lipopolysaccharide-stimulated neutrophils [62,63]. Clinical trials targeting the IL-17 pathways in patients with moderate to SA have evaluated brodalumab (anti-IL-17-receptor antibody), CJM112 (anti-IL-17A antibody), and risankizumab (anti-IL-23 antibody) but have not demonstrated therapeutic benefits [36,37,64]. However, researchers continue to explore other potential therapies aimed at blocking this pathway, including small-molecule inhibitors and DNAzymes that target the ROR pathways [65].
Therapies targeting macrophages
Macrophages play crucial roles in maintaining immune homeostasis and have various functions related to host defense and resolution of inflammation. However, analysis of transcriptomic profiles suggests that macrophages may be functionally impaired in patients with asthma [66]. When exposed to lipopolysaccharide or IFN-γ stimuli, polarization produces M1 macrophages, which are associated with proinflammatory responses. These macrophages release proinflammatory cytokines such as IL-1β, TNF-α, and IL-6, resulting in an influx of neutrophils into the airways and a diminished response to standard anti-inflammatory treatments [67]. Therefore, when developing targeted therapies for patients with T2L, the roles of macrophages and their cytokines must be considered.
The proinflammatory cytokine IL-1β plays an important role in NA. The release of IL-1β from macrophages is regulated by intracellular complexes called inflammasomes. The activation of inflammasomes leads to the production of active IL-1β, which maintains the inflammatory response in patients with NA [67]. Studies have reported correlations between IL-1β, the nucleotide-binding domain leucine-rich repeat pyrin domain-containing (NLRP)3 inflammasome, asthma severity, and the presence of neutrophilic airway inflammation. In addition, elevated levels of IL-1β are positively correlated with the concentration of extracellular DNA in patients with SA. Furthermore, administration of recombinant IL-1β stimulates the recruitment of neutrophils, macrophages, and lymphocytes to the airways; this also increases AHR to methacholine in mice, suggesting the involvement of IL-1β in the recruitment of immune cells and the formation of NETs in patients with NA [68,69].
Therapeutic strategies that target the IL-1β signaling pathways are also being investigated. In a phase 1 clinical trial with healthy subjects, the IL-1 receptor antagonist anakinra reduced airway neutrophilia and increased the levels of IL-1β, IL-6, and IL-8 in response to lipopolysaccharide [39]. In a small study on patients with asthma, canakinumab, a monoclonal antibody that targets IL-1β, reduced the late asthmatic response and decreased IL-1β levels [40]. Recent research on asthma has highlighted the function of bacterial extracellular vesicles [70–75], and vesicles derived from Micrococcus luteus may be used to target neutrophilic airway inflammation. These extracellular vesicles can inhibit both the activation of ILC3 and the production of IL-1β in monocytes by inactivating the NLRP3 inflammasome [76]. Further studies are needed to identify additional biomarkers associated with elevated IL-1β levels in patients with NA and to identify novel biologics that target this pathway.
The multifunctional cytokine IL-6 has been implicated in the pathogenesis of asthma and is associated with airway dysfunction. Studies have reported increased levels of IL-6 in the serum [77], sputum [78], and plasma [79] of patients with asthma, compared to healthy individuals. These increased levels are correlated with increased asthma severity, decreased lung function (FEV1%), obesity, metabolic dysfunction, and elevated neutrophil counts in the blood. IL-6 exacerbates chronic airway inflammation by influencing the balance between regulatory T cells and Th17 cells, resulting in the recruitment of myeloid cells to inflamed tissue and contributing to fibrosis [80]. Moreover, in obese patients with asthma, elevated IL-6 levels are associated with deficiencies in airway cytotoxic CD8+ T cells, particularly in patients with T2L [81]. The role of IL-6 in NA should be investigated in randomized controlled trials that evaluate the efficacy of IL-6 signaling inhibitors in patients with SA. One example of a drug that targets IL-6 is sirukumab, a fully human monoclonal antibody used to treat rheumatoid arthritis. However, sirukumab has been withdrawn due to safety concerns that include a potentially increased risk of mortality [41,82]. A phase 2 clinical trial is currently assessing the efficacy and safety of the IL-6 antagonist clazakizumab in patients with moderate to SA [42]. However, to assess the role of IL-6 in NA, patients would need to be selected based on their phenotypes.
TNF-α has been implicated in the pathophysiology of NA. Previous studies have shown that inhalation of TNF-α increases AHR and sputum neutrophilia in healthy individuals [83]. In addition, in patients with refractory SA, isolated peripheral blood monocytes contain increased levels of membrane- bound TNF-α receptor 1 and TNF-α-converting enzyme [43]. This suggests that there is a relationship between dysregulation of TNF-α signaling and NA. Consequently, the soluble TNF-α receptor blocker etanercept was tested in a small study involving patients with refractory SA [43]. Treatment with etanercept can improve asthma-related quality of life and PC20 (a measure of AHR). However, the results of using TNF-α antagonists, such as golimumab, in patients with severe uncontrolled asthma have been inconsistent. Some studies have not found significant differences in lung function (FEV1%) or the number of asthma exacerbations between treatment groups who received golimumab and placebo groups. Importantly, golimumab is also associated with serious side effects, including infections and malignancies [44]. Clearly, TNF-α has complex roles in the pathophysiology of asthma, and further studies are needed to evaluate the potential risks and benefits of using TNF-α antagonists to manage severe NA.
PGA: MECHANISMS AND THERAPEUTIC TARGETS
The pathophysiological processes underlying PGA are not completely understood. The lack of eosinophils and neutrophils in the airways suggests that PGA may have different underlying mechanisms from NA and MGA. Indeed, structural remodeling of the airway may be the key pathophysiological mechanism in PGA [27]. Transforming growth factor (TGF)-β signaling pathways can promote collagen deposition, myofibroblast proliferation, and subepithelial fibrosis, and these pathways may stimulate airway remodeling in patients with PGA [84]. Epithelial–mesenchymal transitions, which involve epithelial cells acquiring mesenchymal characteristics, may also contribute to airway remodeling in patients with PGA. Inhibiting the functions of proteins such as Smad3 or connective tissue growth factor can attenuate airway remodeling by inhibiting TGF-β signaling and collagen deposition [85]. In addition, TGF-β receptor blockers, such as galunisertib, have been found to restrict fibrosis in preclinical models and clinical trials that have investigated various fibrotic diseases [45]. Another potential therapeutic option is pirfenidone, an antifibrotic agent that can reduce collagen deposition and fibrosis in several lung diseases [46]. Further studies are needed to evaluate the potential of such inhibitors to alleviate TGF-β-mediated airway remodeling in patients with PGA.
CONCLUSIONS
T2L can be subdivided into three phenotypes: PGA, NA, and MGA. Each of these is associated with characteristic pathophysiological mechanisms and therapeutic challenges. Advancing our understanding of these phenotypes and developing suitable targeted therapeutic strategies are crucial for improving outcomes in patients with T2L. More studies are needed to unravel the underlying mechanisms associated with each phenotype and to identify specific biomarkers that can facilitate disease diagnoses and monitoring. In this way, clinicians can achieve better disease control in patients with T2L.
Notes
CRedit authorship contributions
Quang Luu Quoc: writing - original draft; Youngwoo Choi: writing - original draft; Gyu-Young Hur: writing - original draft; Hae-Sim Park: writing - review & editing, visualization, supervision, project administration, funding acquisition
Conflicts of interest
The authors disclose no conflicts.
Funding
This research was supported by the Korea Health Technology R&D Project with a Korea Health Industry Development Institute (KHIDI) grant, funded by the Ministry of Health and Welfare, Republic of Korea (Grant No. HR16C0001).