Distinct functions of eosinophils in severe asthma with type 2 phenotype: clinical implications
Article information

Abstract
Asthma is commonly recognized as a heterogeneous condition with a complex pathophysiology. With advances in the development of multiple medications for patients with asthma, most asthma symptoms are well managed. Nevertheless, 5% to 10% of adult asthmatic patients (called severe asthma) are in uncontrolled or partially controlled status despite intensive treatment. Especially, severe eosinophilic asthma is one of the severe asthma phenotypes characterized by eosinophilia in sputum/blood driven by type 2 immune responses. Eosinophils have been widely accepted as a central effector cell in the lungs. Some evidence has demonstrated that persistent eosinophilia in upper and lower airway mucosa contributes to asthma severity by producing various mediators including cytokines, chemokines and granule proteins. Moreover, extracellular traps released from eosinophils have been revealed to enhance type 2 inflammation in patients with severe asthma. These novel molecules have the ability to induce airway inf lammation and hyperresponsiveness through enhancing innate and type 2 immune responses. In this review, we highlight recent insight into the function of eosinophil extracellular traps in patients with severe asthma. In addition, the role of eosinophil extracellular vesicles in severe asthma is also proposed. Finally, current biologics are suggested as a potential strategy for effective management of severe eosinophilic asthma.
INTRODUCTION
Asthma is a complicated inflammatory disease in the lower airways presenting diverse pathophysiological characteristics [1]. To understand the key features of asthma, several studies have attempted to classify patients according to asthma phenotypes (clinical presentations) and endotypes (molecular pathways) [2,3]. Asthma was once divided into non-atopic (intrinsic) and atopic (extrinsic) asthma; however, this classification had limitations in distinguishing between groups [4]. Recently, asthma has commonly been classified as type 2 (eosinophilic) or nontype 2 (non-eosinophilic) phenotype based on their biological mechanisms [5,6]. Although the identification of asthma subtypes based on clinical, functional and molecular parameters becomes mandatory in the management of asthma, 5% to 10% of the adult asthmatics still remain refractory to current medications [7]. Among them, some are suffering from more severe asthma symptoms and frequent exacerbations with poor quality of life due to local and systemic eosinophilia [8]. Emerging evidence has revealed the importance of eosinophils in both pathogenesis and treatment of severe asthma [9]. This review summarizes (1) the characteristics of severe asthma based on phenotypes and endotypes, (2) the distinct function of eosinophils, and (3) current biologics for better symptom control in severe eosinophilic asthma.
CHARACTERISTICS OF SEVERE ASTHMA
To date, several studies have deepened our understanding of the clinical characteristics of severe asthma. Most patients with severe asthma have consistent features such as frequent/severe asthma exacerbations and progressive lung function decline [10-12], require high-dose inhaled corticosteroids (ICSs) with an additional controller and/or systemic corticosteroids, but their symptoms are not fully controlled with currently available medications [13,14]. Severe asthma is composed of diverse phenotypes according to distinct pathophysiological processes; however, these phenotypes overlap in terms of clinical/physiological outcome and response to treatment [15]. Severe eosinophilic asthma is different from non-severe eosinophilic asthma, although these 2 phenotypes share similar characteristics of eosinophilia in asthmatic airways [16]. It is shown that patients with severe eosinophilic asthma are older, and present higher peripheral/airway eosinophilia, higher fractional exhaled nitric oxide levels and frequent exacerbations, whereas those with non-severe eosinophilic asthma are younger and present higher serum total/specific IgE levels which can be suppressed by anti-inflammatory agents [17-19]. Moreover, persistent airflow limitation and higher prevalence of upper airway pathologies such as chronic rhinosinusitis (CRS)/nasal polyposis (NPs) (with mucosal eosinophilia), are commonly noted [20]. In addition, there is a special phenotype of severe eosinophilic asthma called aspirin-exacerbated respiratory disease (AERD) which is characterized by (1) nonsteroidal anti-inflammatory drug hypersensitivity, (2) moderate to severe persistent asthma, and (3) higher prevalence of CRS/NPs (where intense eosinophilia is noted in upper and lower airway mucosa) and commonly found in middle-aged females. Major pathogenic mechanisms are activated type 2 responses/eosinophils and overproduction of cysteinyl leukotrienes [21]. Furthermore, recent studies have highlighted the role of epithelial cells interacting with eosinophils (via activated surfactant protein D or folliculin) [22-25]. Therefore, further understandings about distinct functions of eosinophils may provide the right targets and biologics in the management of severe eosinophilic asthma.
CLINICAL SIGNIFICANCE OF EOSINOPHILS IN SEVERE ASTHMA
Eosinophils have been highlighted as the hallmark of severe eosinophilic asthma. They are major effector cells contributing to the pathogenesis of asthma by inducing type 2 inflammation and airway hyperresponsiveness (AHR) [26]. It has also been well demonstrated that eosinophils enhance type 2 immune responses by releasing several molecules such as cytokines, chemokines and granule proteins in response to parasitic helminth, bacterial, fungal and viral infection as well as allergens [27]. Persistent airway inflammation induced by eosinophils leads to constant tissue damage, resulting in smooth muscle thickening, goblet cell hyperplasia and extracellular matrix protein deposition called airway remodeling [28]. In inflammatory conditions, eosinophils produce cytokines (interleukin 2 [IL-2], IL-4, IL-5, IL-10, IL-12, IL-13, IL-16, IL-18, tumor necrosis factor-α [TNF-α], and transforming growth factor-α/β), chemokines (macrophage inflammatory protein 1 alpha [MIP-1α], regulated upon activation, normal T cell expressed and secreted [RANTES], and eotaxin-1) and other factors (vascular endothelial cell growth factor and metalloproteinases), and release granule proteins including major basic protein (MBP), eosinophil cationic protein (ECP), eosinophil peroxidase (EPO), and eosinophil-derived neurotoxin (EDN) which were proven to damage airway tissues in various ways [29]. Our recent study demonstrated significantly elevated levels of serum EDN in patients with severe asthma [30], suggesting that EDN, an indicator of eosinophils degranulation, is closely associated with asthma severity. In addition, emerging evidence has revealed that activated eosinophils produce novel molecules, such as extracellular traps or extracellular vesicles (EVs), which will be discussed in the following paragraphs.
Interactions between eosinophils and other immune cells exacerbate asthma symptoms (Fig. 1). Eosinophils certainly respond to IL-5 produced by T cells [31-33]. In the lungs, T cells are the main source of IL-5, which is critical for the recruitment, proliferation, survival and activation of eosinophils. Moreover, neutrophils are involved in eosinophil stimulation to induce airway inflammation by producing extracellular traps in severe asthma [34]. An important role of neutrophils interacting with eosinophils has also been suggested in occupational asthma [35]. Dendritic cell activation and migration could be promoted by EDN released from eosinophils [36]. Furthermore, eosinophils extensively communicate with tissue-resident mast cells [37]. Thus, activation, degranulation, interaction, survival and migration of eosinophils should be suppressed for the management of patients with severe asthma.
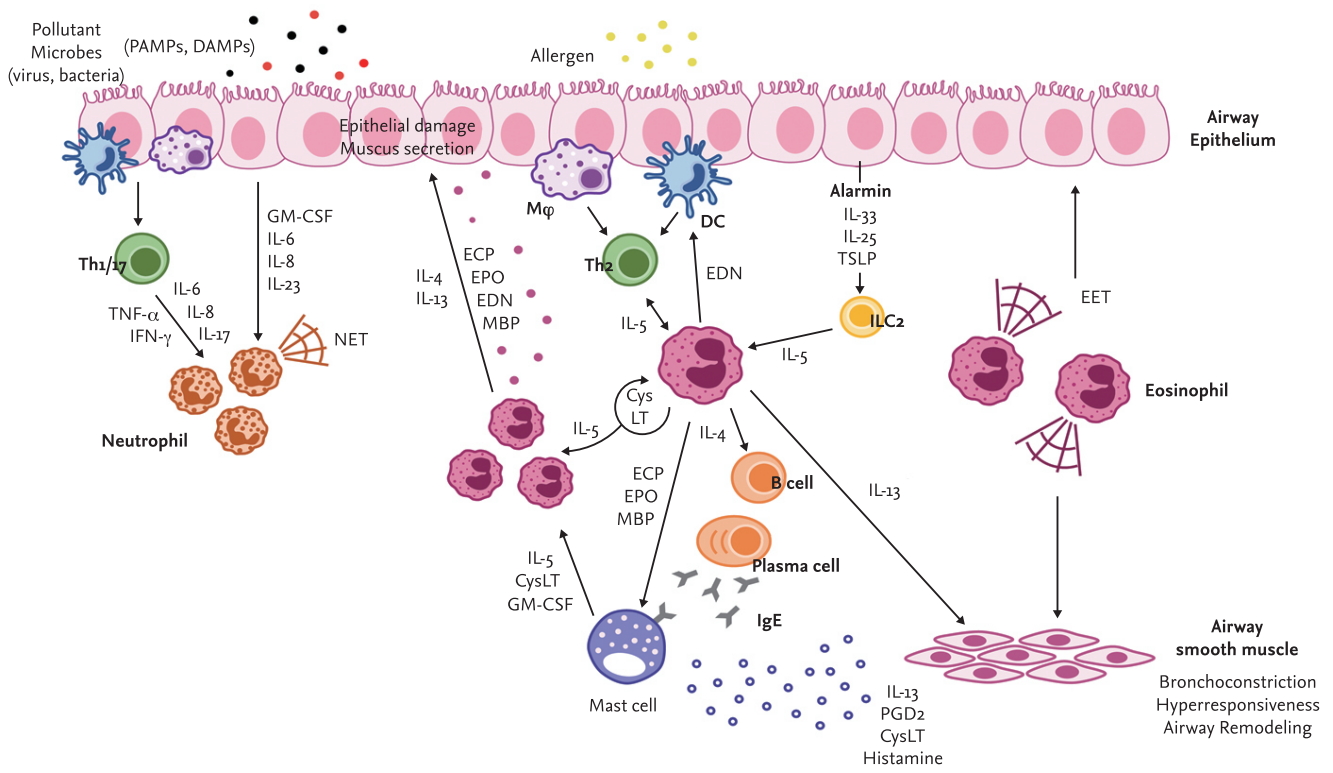
Interactions between eosinophils and various immune cells contributing to airway inflammation in severe asthma with a type 2 phenotype. PAMP, pathogen-associated molecular pattern; DAMP, damage-associated molecular pattern; Th1/2/17, type 1/2/17 T helper cells; ILC2, type 2 innate lymphoid cell; Mϕ, macrophage; DC, dendritic cell; GM-CSF, granulocyte-macrophage colony-stimulating factor; ECP, eosinophil cationic protein; EPO, eosinophil peroxidase; EDN, eosinophil-derived neurotoxin; TSLP, thymic stromal lymphopoietin; NET, neutrophil extracellular traps; EET, eosinophil extracellular traps; CysLT, cysteinyl leukotriene; PGD2, prostaglandin D2; TNF-α, tumor necrosis factor-α; IFN-γ, interferon-gamma; IL, interleukin.
ROLE OF EOSINOPHIL EXTRACELLULAR TRAPS IN SEVERE ASTHMA
Immune function (innate and adaptive immunity) is classically defined as a host defense to recognize and eliminate pathogens. In innate immunity, neutrophils have been intensively studied as the first line of barrier against pathogen invasion. Neutrophil phagocytosis is a well characterized innate immune mechanism; however, an unexpected phenomenon has been also observed that neutrophil-forming extracellular DNA fibers bind to pathogens during host defense. These web-like chromatin structures were firstly found in neutrophils and termed neutrophil extracellular traps (NETs) [38]. NETs have been demonstrated to be implicated in multiple diseases along with several immune-modulatory functions [34,39,40]. Similar to neutrophils, extracellular traps have subsequently been observed from other cells including mast cells, monocytes, macrophages, and eosinophils [41-44].
Eosinophils release web-like chromosomes upon appropriate stimulation. DNA presented in extracellular traps was shown to be of mitochondrial origin, and multiple granule proteins were co-localized in DNA strands [45]. In addition, recent findings have suggested that most extracellular traps are composed of histone-bound DNA that is nuclear in its origins [46]. The formation of eosinophil extracellular traps (EETs) was once thought to be processed by the non-apoptotic cell death pathway [47,48] but these molecules were also found to be released from activated eosinophils independently of cellular cytoskeletal remodeling [49]. Moreover, EET formation was induced by nicotinamide adenine dinucleotide phosphate in an oxidase-dependent manner, which is distinct from apoptosis and necrosis [46].
Although EETs play an important role in innate immunity against extracellular pathogens, they have been related to host tissue damage, contributing to the pathogenesis of allergic diseases [50]. Previous studies have shown that EETs are often associated with blood and tissue eosinophilia [45,51]. Harmful effects of EETs on the airways of patients with asthma have also been demonstrated [44,47]. Furthermore, the proportion of eosinophils releasing extracellular traps was more elevated under the condition of severe airway inflammation [52,53]. Although the mechanisms by which EETs disturb immune responses have not been fully understood, our recent study demonstrated that peripheral EET-forming eosinophil and group 2 innate lymphoid cell (ILC2) counts are elevated in severe asthmatics compared to non-severe asthmatics, with a positive correlation between them and higher levels of peripheral/lung IL-33 and thymic stromal lymphopoietin (TSLP) [54,55]. An in vivo experiment showed that EETs could activate ILC2s in lung tissues through stimulation of airway epithelium to produce IL-33 and TSLP [54], which was attenuated by anti-IL-33 antibody treatment, suggesting that EETs play a crucial role in perpetuating type 2 airway inflammation in severe eosinophilic asthma. These findings suggest that biologics targeting epithelial cytokines may be beneficial in patients with severe eosinophilic asthma (with steroid resistance) via suppressive effects of the EET-ILC axis.
EOSINOPHIL EXTRACELLULAR VESICLES IN SEVERE ASTHMA
In the past, EVs were thought to be cell debris, but now it is certain that they are important mediators produced by cellular processes [56]. EVs are small membranous particles made up of lipid bilayers that contain biological information. Indeed, EVs are composed of a wide spectrum of molecules such as lipids, proteins, and nucleic acids. In terms of a heterogeneous collection of membrane-bound carriers, the function of EVs in cell-to-cell communication has been emphasized [57]. In addition, accumulating evidence supports that EVs are involved in pathophysiological processes of chronic inflammatory diseases such as cancer, metabolic disorders, and allergic disease [58-61]. EVs can promote airway inflammation through regulating recruitment, activation, and differentiation of immune cells and structural cells. Although every cell secretes EVs, especially eosinophils from patients with asthma have been shown to release larger amounts of EVs compared to those released from eosinophils of healthy subjects. The higher levels of EVs in asthmatic patients could lead to more serious symptoms when the EVs are stimulated to release their contents [62,63]. Furthermore, EV production was increased when eosinophils were stimulated with eotaxin-1 or TNF-α [64]. EVs derived from eosinophils contain the components of granule proteins such as MBP, ECP, and EPO; therefore, they similarly contribute to the pathogenesis of asthma. Moreover, EVs released from patients with asthma have been demonstrated to enhance eosinophil migration by up-regulating the expression of adhesion molecules [63]. A recent study has suggested that EVs drive the progression of severe asthma [65]. Diverse miRNAs in EVs have been proposed to be associated with asthma severity [66]. Despite growing interest, the exact mechanism of EVs in the pathogenesis of asthma or any applicable therapy has not yet been found. Further studies are needed to understand the role of eosinophil-derived EVs, which enables us to understand the complicated functions of eosinophils in asthmatic airways. It is suggested that EVs derived from eosinophils may be a potential biomarker for diagnosing asthma and classifying its phenotypes, especially severe eosinophilic asthma.
MANAGEMENT OF SEVERE ASTHMA
According to the Global Initiative for Asthma 2019 guidelines, severe asthma is defined as uncontrolled asthma despite proper adherence to optimized step 4/5 therapy and treatment of contributory factors, or asthma which worsens when doses of anti-asthmatic medications are decreased [67]. As severe asthma is associated with significant morbidity and mortality, several medications have been developed and used (Table 1). Conventionally, ICSs with long-acting beta-agonists (LABAs) are regarded as the first-line therapy for most patients with severe asthma [68]. In addition, systemic corticosteroids can often be administered as an add-on therapy to prevent asthma exacerbation [11,13]. ICSs, known as glucocorticoids, are known to directly or indirectly suppress various immune/structural cells and cytokines involved in airway inflammation [69]. At the gene expression level, they increase or decrease various transcription factors related to airway inflammation [70]. They could increase anti-inflammatory cytokines as well as decrease inflammatory cytokines, chemokines, inflammatory enzymes and adhesion molecules. At the cellular level, corticosteroids inhibit survival or recruitment of various inflammatory cells (such as eosinophils, T cells, and mast cells) and structural cells including epithelial cells in asthmatic airways [71]. Thus, ICS treatment could reduce the number of airway eosinophils and the recovery of epithelial cell injury, improving AHR/lung functions [72,73], and reducing asthma exacerbations [74]. Although an anti-inflammatory effect of corticosteroids is widely accepted, their use in clinical practice is still limited because adverse effects of corticosteroids (in a high-dose or long-term usage of systemic steroids) and decreased responsiveness to corticosteroids (insensitivity or steroid-dependence) have been found in some patients with severe eosinophilic asthma [12,75,76]. Also, since the dose-response curve of ICSs is flat, several addon therapies need to be included for the management of patients with severe asthma who are not effectively controlled with conventional anti-inflammatory medications such as medium-to-high doses of ICSs-LABAs and additional anti-leukotrienes (LTRAs) [77]. Several studies have shown that additional use of LABAs with ICSs is more effective than escalating the dose of ICSs in improving lung function and symptoms control [78] and in reducing the frequency of asthma exacerbations [79,80]. However, there is the possibility that regular use of LABAs could increase underlying inflammation in asthma, such as delays in eosinophil apoptosis [81] or blockade of apoptosis induced by corticosteroids [82]. LTRAs decrease eosinophil counts in blood and airways by blocking the cysteinyl leukotriene receptor 1 (cysLT1R) [83,84] and reduce exacerbations when combined with ICSs [84]. CysLTs are important pro-inflammatory mediators in asthma via increasing bronchoconstriction, AHR, vascular permeability and inflammatory cell recruitment [85]. Especially, CysLTs play important roles in the survival, maturation and differentiation of eosinophils as well as the release of IL-4, ECP, and EDN. Also, eosinophils are major sources of CysLT by autocrine or paracrine stimulation [86]. Recently, it has been reported that human ILC2s express CysLT receptors; thus, CysLTs are involved in ILC2 activation [87]. Corticosteroids do not effectively inhibit the CysLT synthesis pathway [71]. LTRAs do not completely suppress excessive CysLT release in severe eosinophilic asthma or AERD [88]. It is controversial whether LTRAs have an anti-inflammatory effect on eosinophils and ILC2s especially in relation to EETs, granule proteins, cytokines, and mediators. Therefore, there are unmet needs to develop alternative or additional medications for better control of eosinophils and severe asthma.
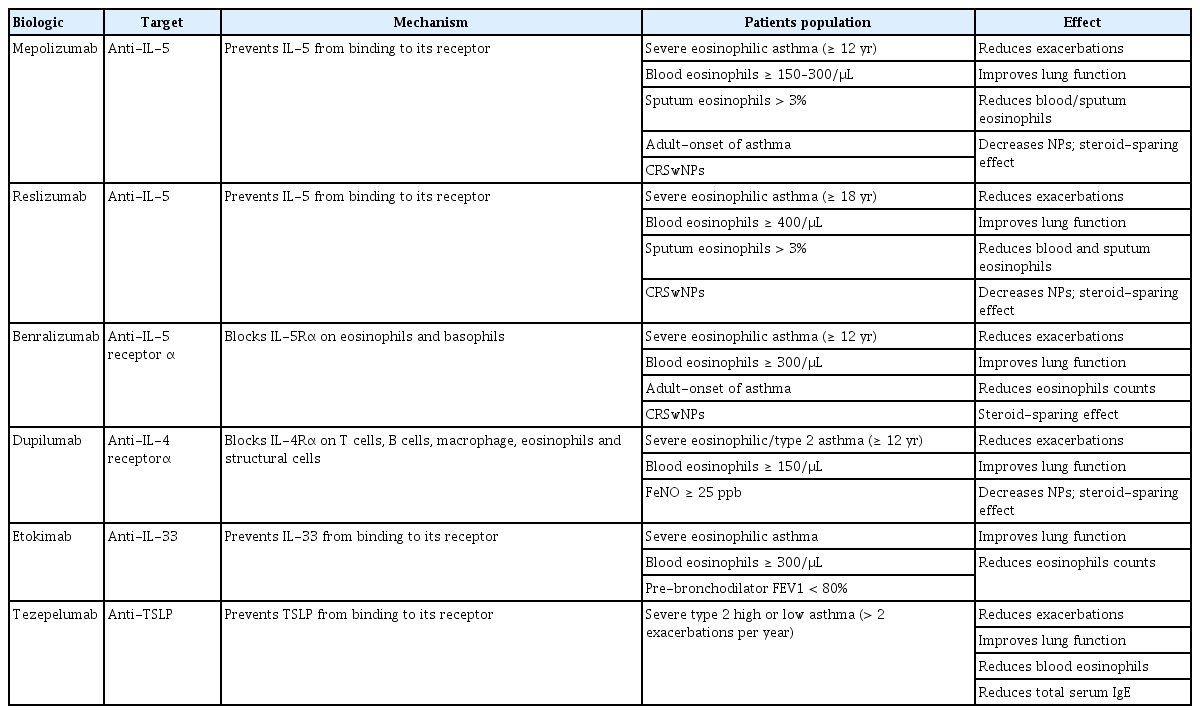
Various medications developed to manage asthma severity
Many studies have been attempted to block the interaction between IL-5 and its receptor in eosinophilic inflammation because IL-5 is a key cytokine involved in eosinophil growth, maturation, activation and survival. Benralizumab is a monoclonal antibody against the alpha subunit of the IL-5 receptor. This antibody provides a potential benefit in decreasing exacerbation and improving lung function in uncontrolled severe asthma with elevated blood eosinophil counts by inducing antibody-dependent cell-mediated cytotoxicity [89-91]. Mepolizumab and reslizumab, humanized monoclonal antibodies against IL-5, neutralize circulating IL-5 and decrease the number of eosinophils in sputum and blood [92-95], leading to improvement in lung function in patients with severe eosinophilic asthma. IL-4 has also been suggested to play an important role in the differentiation and proliferation of type 2 helper T cells and B cells [96,97]. Dupilumab, a human monoclonal antibody against the alpha subunit of the IL-4 receptor (an overlapping receptor of IL-4 and IL-13), is regarded as a therapeutic agent of disease mediated by type 2 helper T cells. When this antibody was administered to patients with persistently elevated eosinophil levels, a significant decline in the frequency of severe exacerbation was observed [98,99]. Recently, our data has shown that anti-IL-33 antibody reduces AHR and decreases type 2 cytokine levels in an EET-induced inflammation model in vivo [54]. Considering the critical role of EETs in the pathogenesis of severe eosinophilic asthma, biologics targeting epithelial cytokines, especially IL-33, may provide a potential benefit.
Eosinophilic inflammation is strongly associated with type 2 cytokines, as well as allergen-specific IgE. In addition, alarmin-like cytokines such as IL-33 and TSLP, which are mainly released from airway epithelium, are involved in the development of severe asthma [55,100]. IL-33 activates myeloid and lymphoid innate cells to exacerbate airway inflammation but stimulates eosinophils as well [101]. TSLP is regarded as an IL-7-like cytokine known to be important for inducing type 2 cytokine production, leading to the activation of eosinophils [102]. In a previous study, the efficacy of human monoclonal anti-TSLP antibody in patients with allergic asthma was tested; a potential benefit in attenuating airway inflammation was suggested [103]. Moreover, the effect of anti-IL-33 or anti-TSLP antibody on reduction in AHR was demonstrated in vivo models [54]. Although antibodies against IL-33 and TSLP are under clinical trial, they can be a promising treatment for patients with severe type 2 asthma.
Although various pharmacotherapy and biologics have been approved for the management of severe asthma, there remain unresolved issues about selecting proper targets and patients for effective treatment, depending on its phenotypes/endotypes. Further studies are needed to find potential biomarkers for various phenotypes and endotypes to implement precision medicine.
CONCLUSIONS
Accumulating evidence strongly supports heterogeneity in severe asthma with distinct subtypes. Especially, an important role of eosinophils in type 2 severe asthma has been widely accepted as sputum/blood eosinophilia is associated with more severe symptoms, more frequent exacerbations and lower response to anti-inflammatory medications. Recent studies suggest that novel molecules, including extracellular traps and vesicles released from eosinophils, could enhance type 2 immune responses interacting with airway epithelium in the pathogenesis of severe eosinophilic asthma. However, current anti-asthmatic medications have limitations in completely controlling severe eosinophilic asthma; instead, new biologics targeting eosinophils or epithelial cells can provide potential benefits with some limitations. The development of effective biologics in terms of eosinophil function is essential for better management of severe asthma.
Notes
No potential conflict of interest relevant to this article was reported.
Acknowledgements
This study was supported by a grant from the Korean Health Technology R & D Project, Ministry of Health and Welfare, Republic of Korea (HI16C0992).