


INTRODUCTION
Stimulation of Na+-K+ pump activity by the thyroid hormone in several tissues, including liver, kidney, skeletal muscle and heart has been demonstrated in a wide variety of experimental studies1–10) and has been proposed as a major mediator, i.e. metabolic pacemaker of thyroid thermogenesis11–13). However, despite these observations, the mechanism by which the enhancement of Na+-K+ pump activity is mediated by the thyroid hormone has not been resolved. Although it has been postulated that the effect of thyroid hormone on active cation transport is mediated by direct stimulation of sodium-potassium pump unit synthesis5,7,12), it is not clear how an induction of additional Na+-K+ pump units per se could explain a steady state enhancement of active sodium and potassium transport in the absence of an augementation of passive fluxes of cations opposite to the direction of their active transport.14) Recent evidence does in fact exist to suggest that thyroid hormone increases monovalent cation permeability in isolated rat skeletal muscle15) and perfused rat liver.5,14)
In contrast to the above animal data, little information on the state of the Na+-K+ pump in human patients with altered thyroid function is available; moreover, the available data on human patients appears contradictory to the results in animals. Thus, Cole et al. reported that the Na+-K+ ATPase activity in human erythrocyte membranes was reduced in hyperthyroidism and this finding was consistent with the increase intracellular sodium concentration [Na+]i.16) To further examine this apparent change in the cation transport in red blood cells, we have measured the Na+-K+ pump activity as well as Na+-K+ cotransport (CoT), Na+-Li+ countertransport (CTT) and Na+ passive permeability in patients with hyper- and hypothyroidism.
SUBJECTS AND METHODS
1. Subjects
Nineteen untreated hyperthyroid patients, twelve treated hyperthyroid patients and nine hypothyroid patients with T4 replacement were recruited for this study; the majority were outpatients from the Endocrine Clinic of Kangnam St. Mary’s Hospital. The relevant clinical and thyroid hormone data for each group are shown in Table 1. At the time of the initial measurements, untreated hyperthyroid subjects had been hyperthyroid status for at least 4 weeks, as judged from their clinical history and available laboratory data. The untreated hyperthyroid subjects were not any specific antithyroid drug or other medication, and in particular, no patient was being treated with agents such as corticosteroids which are known to affect the Na+-K+ pump activity. The treated hyperthyroid subjects had been managed with antithyroid drugs for at least 4 weeks. The clinical diagnosis of hypothyroidism was confirmed by the measurement of serum T3, T4 and TSH using standard radioimmunoassays. All of the hypothyroid subjects were managed with T4 replacement.
Thirty-seven control subjects were recruited from the medical student population, nursing personnel and doctors, all of whom were clinically and hormonally euthyroid. The mean age of the normal subjects was 30.2 (range; 24–37 yrs), somewhat younger than that of the untreated hyperthyroid and hypothyroid subjects. In addition, there was approximately equal representation by males and females, in contrast to the preponderance of males in each study group. However, we were unable to detect any age or sex-related differences in the Na+-K+ pump measurement in our control population (Table 1).
2. Methods
1) Preparation of RBC
Venous blood (10 ml) collected in heparinized tubes was centrifuged at 1,750 g for 10 min. The plasma and buffy coat were also aspirated. RBC were then washed twice with isotonic MgCl2 (110 mM). All steps were carried out at 4°C.
2) Determination of Intracellular Na+[Na+]i and K+ [K+]i
Cells were washed 3 times with 10 volumes of ice-cold isotonic MgCl2 solution. After mixing, duplicate hematocrit determinations were made of this suspension. Then 100 μl were diluted 1:50 in deionized water. The concentration of Na+ and K+ of the hemolysate was measured on a Perkin Elmer model 2380 Atomic Absorption spectrophotometer, and expressed as mmol/l of packed red cells by dividing the ionic concentration by the hematocrit and multiplying by the dilution factor.
3) Measurements of Ionic Fluxes
Simultaneous measurements of the Na+-K+ pump, Na+-K+ CoT, Na+-Li+ CTT, and ouabain-and bumetanide-resistant Na+ and K+ fluxes (passive Na+ and K+ permeability) were made according to the modified method of Garay et al.17). Washed red cells were resuspended in cold Mg++ sucrose medium (mM): 75 MgCl2, 85 sucrose, 10 MOPS-Tris (pH 7.4 at 37°C) and 10 glucose, to a hematocrit of 20–25%. A portion of the cell suspension was added (final hematocrit 4–5%) to different cold solutions containing buffered Mg++ sucrose plus the following additions (mM): (1) 2 KCl, (2) 0.1 ouabain, (3) 0.1 ouabain plus 0.02 bumetanide, and (4) 10 LiCl, 0.02 bumetanide, and 0.1 ouabain. At time 0, the tubes were transferred to a 37°C bath for incubation. Sixty minutes after incubation, media 1,2,3, and 4 tubes were transferred to an ice bath for 1 min and then centrifuged at 1,750g for 5 min at 4°C. The supernatant was carefully removed, and the Na+ and K+ concentrations were measured by atomic absorption spectrophotometry, using standards prepared in the appropriate Mg++ sucrose media.
Na+ and K+ effuxes were computed using the following equation:
D Cat. (μmol/l supernatant) is the difference between the external cation concentration (Na+ or K+) after incubation at 37°C and that at 0 time. The Na+-K+ pump activity was calculated by subtracting the Na+ efflux in the presence (medium 2) from that in the absence (medium 1) of ouabain. The Na+-K+ CoT fluxes were obtained by subtracting the Na+ efflux in the presence of ouabain plus bumetanide (medium 3) from that in the presence of ouabain alone (medium 2), while for the calculation of Na+-Li+ CTT, the Na+ efflux in medium 3 was subtracted from that in medium 4. The Na+ fluxes in medium 3 were taken as the ouabain-and bumetanide-resistant Na+ fluxes. The efflux rate constants for ouabain-sensitive and bumetanide-sensitive Na+ effluxes, and the influx rate constant for passive Na+ permeability expressed in h−1, were calculated as the ratio of the respective fluxes to the erythrocyte Na+ content.
RESULTS
1) Intracellular Na+ and K+ Concentration
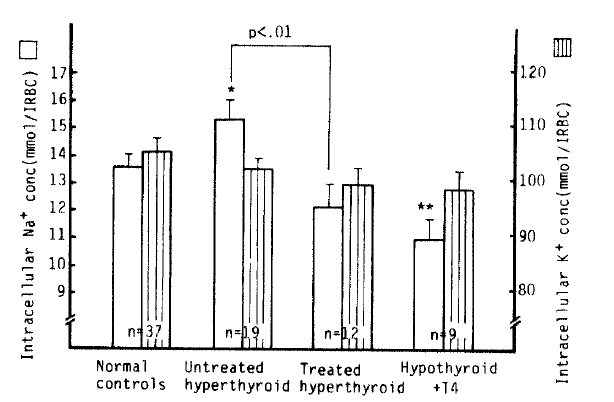
The mean [Na+]i value in the untreated hyperthyroidism group (15.3 ± 0.7 mmol/l RBC) was significantly higher than that in normal subjects (13.6 ± 0.4 mmol/l RBC) (p<.05), but not significantly different from that in the treated hyperthyroidism group (12.1 ± 0.8 mmol/l RBC). The mean [Na+]i value in the hypothyroidism with T4 replacement group (10.9 ± 0.7 mmol/l RBC), however, was statistically significantly lower than that in the control group (p<.01). Also, a statistically significant reduction of [Na+]i values in the treated hyperthyroidism group vs. that of those in untreated hyperthyroidism groups was noted (p<.01). The mean [K+]i values in each group were not significantly different from that of normal subjects (105.8 ± 2.1 mmol/l RBC) (Table 2, Fig 1).
2) Na+-K+ pump (Ouabain-sensitive Na+ efflux)
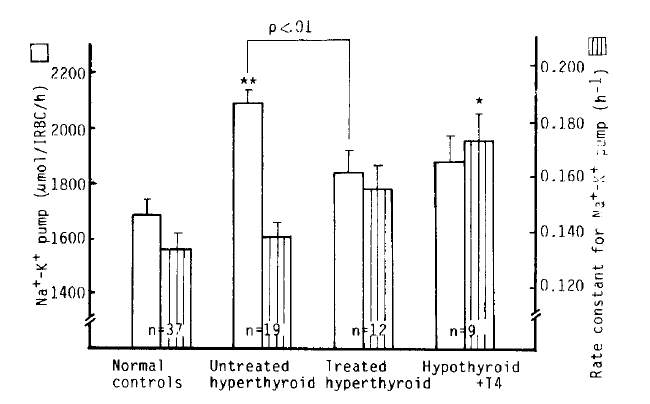
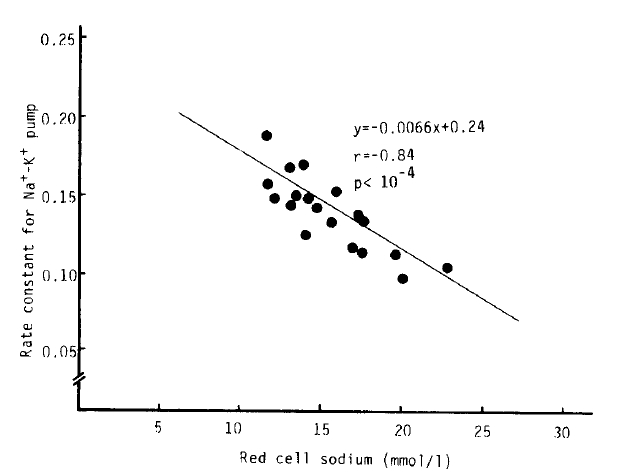
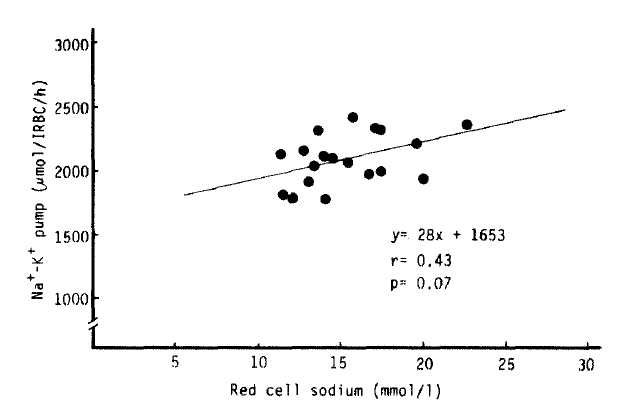
Na+-K+ pump activity in the untreated hyperthyroidism group (2,083 ± 45 umol/l RBC/h) was significantly higher than that of the normal group (1,688 ± 52 umol/l RBC/h) (p<.10−5), but no significant difference was noted from that in the treated hyperthyroidism group and hypothyroidism with T4 replacement group. We observed a significant decrease of Na+-K+ pump activitiy in the patients being treated for hyperthyroidism compared to that of the untreated hyperthyroidism group (1,837 ± 73 vs. 2083 ± 45 umol/l RBC/h, p<.01). The rate constant for ouabain-sensitive Na+ efflux in the hypothyroidism with T4 replacement group (0.1738 ± 0.0095 h−1) was markedly higher than that in the normal group (0.1359 ± 0.0059 h−1) (p<.01). But there was no statistical significance in the rate constant between the normal and untreated hyperthyroidism groups (0.1396 ± 0.0054 h−1). We observed a significant inverse correlation (r = −0.84, p<10−4) between the variation of the rate constant for the ouabain-sensitive Na+ efflux and [Na+]i, but not between the Na+-K+ pump activity and [Na+]i (r = 0.43, p = 0.07) (Table 2, Fig 2,3,4).
3) Na+-K+ Cotransport (Bumetanide-sensitive Na+ efflux)
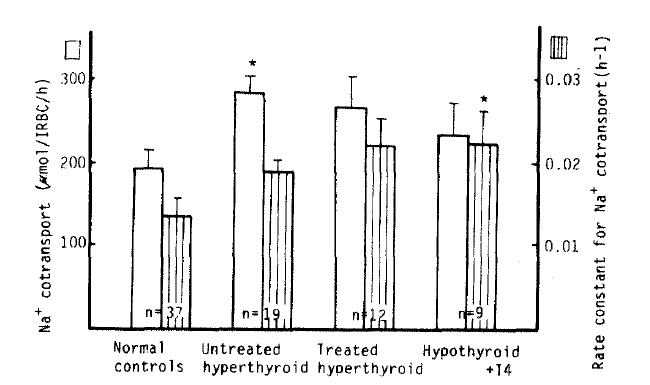
We observed a significant increase of the Na+ CoT value in the untreated hyperthyroidism group versus in the normal subjects (282 ± 20 vs. 192 ± 23 μmol/l RBC/h, p<.05), but no significant difference between the treated hyperthyroidism group and the hypothyroidism with T4 replacement group (266 ± 37 vs. 234 ± 38 μmol/l RBC/h). But, the rate constant for Na+ CoT in the hypothyroidism with T4 replacement group was significantly higher than that in the control group (0.0223 ± 0.0038 vs. 0.0134 ± 0.0022 h−1, p<.05). There was no significant difference between each group in K+ CoT value (Table 2, Fig 5).
4) Na+-Li+ Countertransport (Li+-stimulated Na+ efflux)
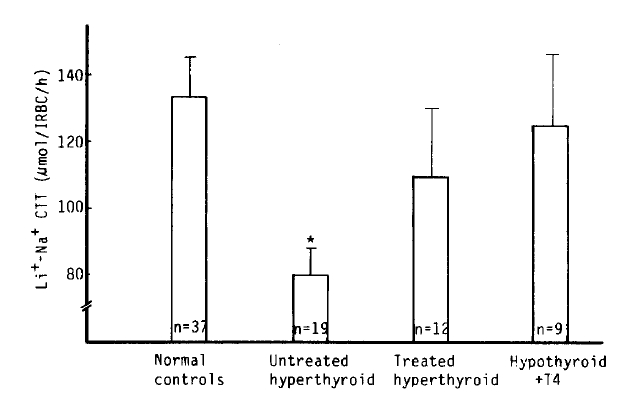
We observed a marked decrease of the Na+-Li+ CTT value in the untreated hyperthyroidism group vs. that of the normal group (80 ± 8 vs. 133 ± 12 μmol/l RBC/h, p<.01). But, there was no significant difference between the treated hyperthypoidism group and hypothyroidism with T4 replacement group (109 ± 21, 125 ± 21 μmol/l RBC/h), (Table 2, Fig 6).
5) Passive Na+ Premeability (Ouabain and bumetanide-resistant Na+ influx)
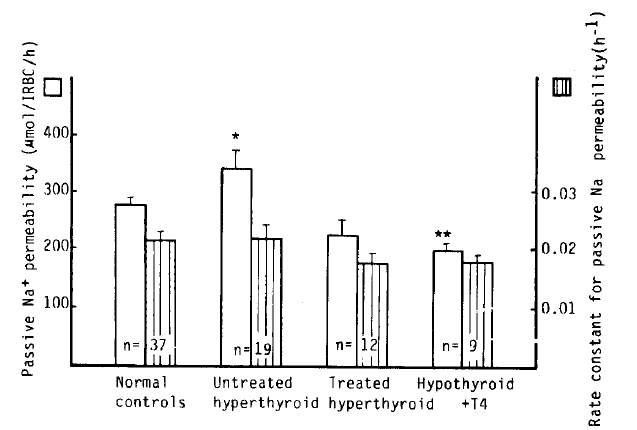
Passive Na+ permeability in patients with untreated hyperthyroidism was markedly higher than that in normal subjects (339 ± 33 vs. 276 ± 11 μmol/l RBC/h, p<.05), whereas that in the hypothyroidism with T4 replacement group was markedly lower than that in normal subjects (198 ± 13 vs. 276 ± 11 μmol/l RBC/h, p<.01). However, the rate constant for passive Na+ permeability was not statistically significant between the groups (Table 2, Fig 7).
DISCUSSION
We have examined the interrelationships between erythrocyte sodium content and sodium transport in a group of healthy subjects and in groups of patients with abnormal thyroid hormones. The sodium content of the erythrocyte changes in a variety of acquired disorders including hypokalemia, digoxin toxicity, hyperthyroidism, hemorrhagic shock, chronic renal failure, and liver disease as well as in hereditary defects of the erythrocyte membrance.18) Boekelman, in 1958, first reported that patients with hyperthyroidism have an elevated concentration of sodium within their erythrocytes.19) Thereafter, many reports subsequently verified this observation and suggested that the determination of red cell sodium might prove useful as a measure of the peripheral action of thyroid hormone.18–22) In this study, we have shown that untreated hyperthyroidism is associated with a significant increase in the intracellular sodium concentration (p<.05); however, treated hyperthyroidism does not bear the same results. In contrast to untreated hyperthyroidism, there was a significant decrease of [Na+]i in patients with treated hyperthyroidism (p<.01). What is the exact mechanism for the significant increase in [Na+]i in untreated hyperthyroidism?
A derangement of ion transport in altered thyroid function has been demonstrated in many tissues.1–10) Sodium movements across human RBC membranes are mediated by several transport systems. The ouabain-sensitive Na+-K+ ATPase is the main mechanism maintaining the Na+ and K+ electrochemical gradients across RBC membranes against the passive Na+ and K+ diffusion. At least three additional transport systems have recently been described in human RBC membrans: (1) Na+-K+ CoT, (2) Na+-Li+ CTT, and (3) passive Na+ permeability.17)
Since Smith and Samuel (1970) reported that the increase in red cell sodium in patients with hyperthyroidism was associated with a decrease in the rate constant for active sodium efflux and significantly higher total active sodium efflux, the arguments for and against these results appeared.20) The thermogenic response to thyroid hormone has been observed in many tissues including heart, kidney, liver and skeletal muscle of rats. Recently, such a thermogenic response has been attributed to the stimulation of energy utilization resulting from the influence of this hormone on the sodium pump activity. Subsequently, thyroid hormone treatment has been shown to augment Na+-K+ ATPase activity, an enzyme system closely related to the sodium pump.1,2,7,12,13,23) Ismail-Beigi and Edelman recently proposed that augmentation of energy expended in active Na+ transport mediates a significant fraction of the thermogenic response to thyroid hormone. In rat liver and skeletal muscle, ouabain-sensitive or Na+-dependent respiration increased significantly in the transitions from the hypothyroid to the euthyroid and from the euthyroid to the hyperthyroid states induced by adminstration of T3.1) Asano et al.4) reported that administration of thyroid hormone induced enhancement of O2-dependent respiration in the liver, kidney and diaphragm of the rat, which was inhibited by cardiac glycoside. So the thermogenesis action of thyroid hormone might be mediated by Na+-K+ ATPase activity. The magnitude of the energy demands of the Na+ pump in the hyperthyroid state determines its importance as a metabolic pacemaker. In the rats studied by Lin and Akera, T3 treatment increased the [3H] ouabain binding site concentration in the liver, kidney, and skeletal muscle but failed to affect it in the brain.9) Although thyroid hormone increased Na+-K+ ATPase activity in many tissues, the result of erythrocyte Na+-K+ ATPase activity in hyperthyroid patients were controversial and little information on the state of the Na+K+ pump in human patients with altered thyroid function is available. However, in human subjects, activity of this enzyme, studied extensively in erythrocytes, has been found to be consistently reduced in the hyperthyoid state. Although Cole and Waddell (1976)16) tried to conclude from their studies that the decreased sodium efflux in the erythrocytes of patients with hyperthyroidism was associated with a decrease in Na+-K+ ATPase activity, the entry number was too small to draw this conclusion. In other studies, the number of Na+-K+ ATPase units in erythrocytes was found to be significantly reduced in patients with hyperthyroidism.18, 22, 24)
Nevertheless, in our study, Na+-K+ pump activity in the untreated hyperthyroidism group was markedly increased (p<.10−5), and in the treated hyperthyroidism group and the hypothyroidism with T4 replacement group, it tended to be higher than normal. Despite increased Na+-K+ pump activity in untreated hyperthyroidism, the rate constant for ouabain-sensitive Na+ efflux was not different from normal, but was significantly higher in the hypothyroidism with T4 replacement group than normal. There was no definite correlations between intracellular Na+ concentration and Na+-K+ pump activity. However, a significant inverse correlation between intracellular Na+ concentration and the rate constant for Na+-K+ pump activity was noted (r = −0.84, p<.10−4).
In this study, Na+-K+ CoT in the untreated hyperthyroid patients was higher than that in the normal patients, and tended to be higher, but not significantly in the treated hyperthyroid patients and the hypothyroidism with T4 replacement group. The exact mechamism for increased Na+-K+ CoT in the untreated hyperthyroid patients was uncertain. The observed increase in the Na+-K+ CoT, however, might be a secondary adaptive response of the cell to maintain normal intracellular ion concentration and transmembrane ion gradients in the face of this enhanced passive Na+ permeability. We found that Na+-Li+ CTT in the untreated hyperthyroid patients was markedly decreased; however, the mechanism was uncertain.
In our study, passive Na+ permeability in the untreated hyperthyroid patients was substantially higher than in the normal group, and tended to be lower in the treated hyperthyroidism and hypothyroidism with T4 group. Haber and Loeb have examined the effect of T3 treatment on the passive efflux of 42K+ from rat liver slices to determine whether an increase in permeability might play a role in the known enhancement of active monovalent cation transport of Na+-K+ ATPase activity induced by thyroid hormone. The magnitude and early onset of the effect of thyroid hormone on cellular potassium efflux raise the possibility that an increase in passive cation permeability may be a proximal event in the mediation of thyroid hormone action.14,15,25) Additional evidence for a thyroid hormone-induced increase in passive monovalent cation permeability has been provided by Folke and Sestoft5) in studies on perfused intact rat liver following T3 treatment in vivo.
As noted by Ismail-Beigi and Edelman, thyroid hormone-dependent activation of the Na+ pump could be a result of the following: (1) a rise in [Na+]i, perhaps as a result of increased membrane permeability to Na+ and consequent stimulation of the Na+ pump; (2) a change in the coupling ratio between the chemical reaction (i.e. ATP split) and transport rate, such that the rate of ATP hydrolysis is increased for a given rate of Na+ transport. As a result there would be a fall in the transmembrane Na+ and K+ gradients; (3) activation of mitochondrial ATP synthesis, resulting in stimulation of the Na+ pump by a local increase in ATP concentration; and (4) direct activation of the Na+ pump. In our study, the first (1) possibility, as either the sole or the predominant mechanism of the mediating role of Na+ transport in thyroid calorigenesis is supported in understanding the status of increased [Na+]i with the enhanced Na+-K+ pump activity in untreated hyperthyroidism.
Then, what is the exact mechanism for the pathologic alterations of cation movement in red blood cells? Abnormal RBC cation movements have been found in a wide variety of clinical states; i.e. (1) genetic abnormalities such as in hereditary spherocytosis; (2) precipitaion of cell contents such as in sickle cell disease, (3) membrane alterations induced by factors extrinsic to the RBC such as in hyperthyroidism and chronic uremia; and (4) formation of negatively charged ion pairs such as in hemolytic anemia.26)
All of the above findings suggest that untreated hyperthyroidism results in an early increase in membrane permeability with an augmentation of passive Na+ influx. The observed increase in the Na+-K+ pump might then be a secondary adaptive response of the cell to maintain normal intracellular ion concentration and transmembrane ion gradients in the face of the enhanced passive Na+ permeability. Although there is now general agreement that thyroid hormone stimulates sodium and potassium transport, the relative importance of this phenomenon in contributing to the thermogenic effect remains uncertain.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print





