


INTRODUCTION
Pancreatic cancer is known to be an aggressive cancer with a 5-year survival rate of 8% [1]. There are various factors related to high aggressiveness, such as aging characteristics, resistance to treatment, and early metastasis. Surgical resection is only curative treatment, but most patients are diagnosed when they are in unresectable state. Moreover, recurrences develop in 75% to 93% of patients, even in those who underwent surgery followed by adjuvant treatment [2,3]. Various molecular changes and responses in tumor and its microenvironment in pancreatic cancer can play a role in invasiveness, growth and chemoresistance, leading to highly aggressive tumor behavior. However, the mechanism of early metastasis in pancreatic cancer is not yet known.
Wnt/╬▓-catenin signaling plays an important role in many biological processes, including carcinogenesis and progression. In various solid tumors, activation of Wnt/╬▓-catenin signaling is known to be involved in invasion and metastasis. In colon cancer, it promoted scattering of cancer cells and metastasis by inhibition of the phosphatidylinositide 3-kinases-protein kinase B (PI3K-Akt) signaling pathway [4]. It also regulates metastasis by matrix metallopeptidase 9 (MMP-9) expression in bladder cancer [5]. Wnt/╬▓-catenin activation was observed in 65% of pancreatic adenocarcinomas, associated with invasion and metastasis or invasion [6-8]. Some researchers have suggested that the Wnt signaling pathway regulates migration, invasion, and metastasis in pancreatic cancer by producing pancreatic stellate cells [9]. As reported in previous studies, the Wnt/╬▓-catenin pathway can be directly or indirectly involved in the mechanism of metastasis.
In the present study, we tried to determine the role of the Wnt/╬▓-catenin signaling pathway by using pancreatic cancer cell lines and human tissue.
METHODS
Cell lines
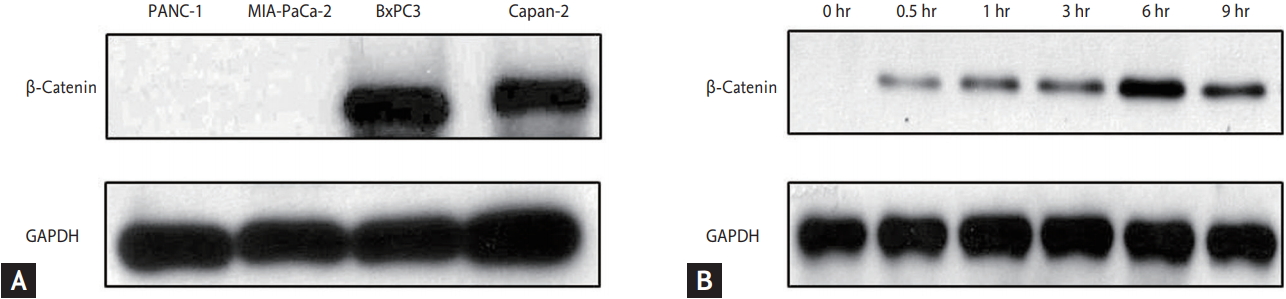
We used pancreatic cancer cell lines including Panc-1, Mia-paca2, BxPC3, Capan2, which were obtained from American Type Culture Collection (ATCC; Manassas, VA, USA). They were maintained in DulbeccoŌĆÖs Modified Eagle Medium (DME; Hyclone, Logan, UT, USA) or Roswell Park Memorial Institute 1640 Medium (RPMI, Hyclone) containing 10% fetal bovine serum (Hyclone) and 100 U/mL penicillin-streptomycin (Hyclone) maintained at 37Ōäā in humidified air containing 5% CO2. We performed the assessment of ╬▓-catenin expression in each cell line (Fig. 1A).
Western blotting assay
Cells were lysed in protein extraction buffer RIPA (Elpis Biotech, Daejeon, Korea) with protein inhibitor cocktail (Roche, Berlin, Germany). Protein quantification was performed using BCA assay (Thermo Scientific, Rockford, IL, USA). Protein (5 to 20 ╬╝g/lane) was separated by 6% to 10% SDS-PAGE and transferred to polyd-vinylidene difluoride membrane (PVDF, GE Healthcare Life Sciences, Freiburg, Germany). Blots were blocked with 5% skim milk solution and incubated with primary antibodies overnight at 4Ōäā. Next day, the blot was incubated with the secondary antibody for 1 hour, and was detected with ECL (Recenttec, Taipei, Taiwan). We used primary antibodies at a specific dilution; glyceraldehyde 3-phosphate dehydrogenase (GAPDH, Abcam, Cambridge, MA, USA) diluted 1:5,000, CD44v6 (Abcam) diluted 1:500, CD44 (GeneTex Inc., Irvine, CA, USA) diluted 1:500, ╬▓-catenin (ab6302, Abcam) diluted 1:4,000, and active-╬▓-catenin (Merk Miillipore, Burlington, MA, USA) diluted 1:300.
Cell invasion assay
Cell invasion assays were conducted on 6.5 mm Transwell with 8.0 ╬╝m pore size polycarbonate membrane insert. For the coating of invasion assay, matrigel (Corning) was diluted at a 1/20 dilution rate with serum-free media and 100 ╬╝L matrigel dilution buffer was coated onto the upper compartment of the cell culture insert. After incubation for 2 hours at 37Ōäā, the cell culture insert was ready for seeding. Pancreatic cancer cells (1.0 ├Ś 105 to 2.0 ├Ś 105) were placed in matrigel-coated upper chambers with serum-free media for invasion assays and a medium containing 10% serum media was used as a chemoattractant in the lower chambers. After incubation at 37Ōäā, invaded cells were stained using DiffQuik staining kit (Sysmex, Kobe, Japan). The images of cells were photographed with EVOS Cell Imaging Systems (Left Technologies, Gaithersburg, MD, USA) at ├Ś200 magnification and the cell number was counted in three random fields of view.
Activation of ╬▓-catenin
Application of Wnt-3a conditioned media
We collected media from Wnt-3a-expressing L cells and cultured media and control media from mouse L cells, prepared according to the manufacturerŌĆÖs instructions (ATCC). We tested the Wnt-3a conditioned media by ╬▓-catenin expression using Western blot. ╬▓-Catenin expression was maximum at 6 hours after Wnt-3a conditioned media (Fig. 1B); therefore, we maintained Wnt-3a stimulation by changing the media every 6 hours.
Transfection of ╬▓-catenin
For stable expression of ╬▓-catenin, we used ╬▓-catenin pcDNA3 (Plasmid 16828#, Addgene, Cambridge, MA, USA), pcDNA3-S33Y ╬▓-catenin (Plasmid #19286, Addgene), the mutant form and pcDNA3 (Invitrogen, USA) as the control vector. The PANC1 cell line, non ╬▓-catenin expressing cell line, was seeded in 6-well dishes and then ╬▓-catenin was transfected using Lipofectamine 2000 (Thermo Fisher, San Jose, CA, USA). After 48 hours, the cells were harvested and Western blotting assays were performed.
Inhibition of ╬▓-catenin
XAV939 treatment
XAV939 (Selleckchem, Houston, TX, USA), a Wnt/╬▓-catenin signaling inhibitor, was diluted to a final concentration of 1 ╬╝M, and then it was applied to the BxPC3 cell line. We harvested co-cultured cells and performed Western blot.
Application of siRNA for ╬▓-catenin
The BxPC3 cell line was cultured in a 6-well plate, and it was treated with 20 nmol/L control siRNA (Sc37007; Santa Cruz) and ╬▓-catenin siRNA (Sc292009; Santa Cruz) by adding TransIT-TKO Transfection Reagent MIR 2150 (Mirus, Madison, WI, USA). The siRNAs were cultured for 48 hours and then cell lysates were subsequently prepared for Western blot to verify knockdown of ╬▓-catenin.
Tissue microarray and immunohistochemical staining
Paraffin blocks with tumor samples were obtained from 73 patients who had undergone curative resection for pancreatic cancer between January, 2001 and December, 2009. Tissue microarray was constructed as mentioned previously [10]. We performed immunohistochemical staining with the following primary antibodies at specified dilutions: Wnt-3a (Abcam) diluted 1:100, Wnt-5a (Abcam) diluted 1:50, and ╬▓-catenin (Abcam) diluted 1:100. The results were interpreted by two independent pathologists who were blinded to the specific diagnosis and prognosis of each case. The staining intensity was scored on a three-tiered scale: score 0, less than 10% of cells positive; 1, 10% to 49% positive; and 2, more than 50% of cells positive. We also reviewed the patientsŌĆÖ medical records to analyze the clinical outcome according to Wnt/╬▓-catenin expression. This study was approved by our Institutional Review of Board: Board in Seoul St. Mary's Hospital (KC11SISI0184).
Statistical analysis
All data are presented as mean ┬▒ standard deviation (SD) of at least three separate experiments. Student t tests were used for transwell invasion assay. The chi-square and Fisher exact tests were used to analyse correlations between immunohistochemical profiles and clinicopathologic variables. Kaplan-Meier tests were used to analyse the survival data. Statistical significance was set at p < 0.05. Statistical analyses were performed using the IBM SPSS version 24.0 (IBM Corp., Armonk, NY, USA).
RESULTS
Cell invasion assay
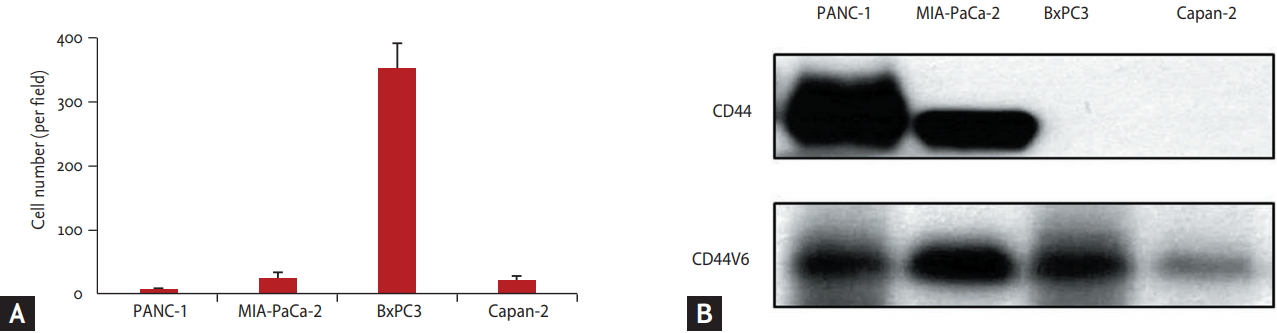
On Transwell invasion assay, the BxPC3 cell line, high ╬▓-catenin expressing cell line was the most invasive, and Panc-1, the non ╬▓-catenin expressing cell line showed weak invasiveness. It was not associated with expression of CD44 or CD44 variant, known as the invasion related protein (Fig. 2). We chose BxPC3 and Panc-1 cell lines for further experiments as ╬▓-catenin expressing and non ╬▓-catenin expressing cell lines, respectively.
Over expression of ╬▓-catenin enhanced cell invasiveness
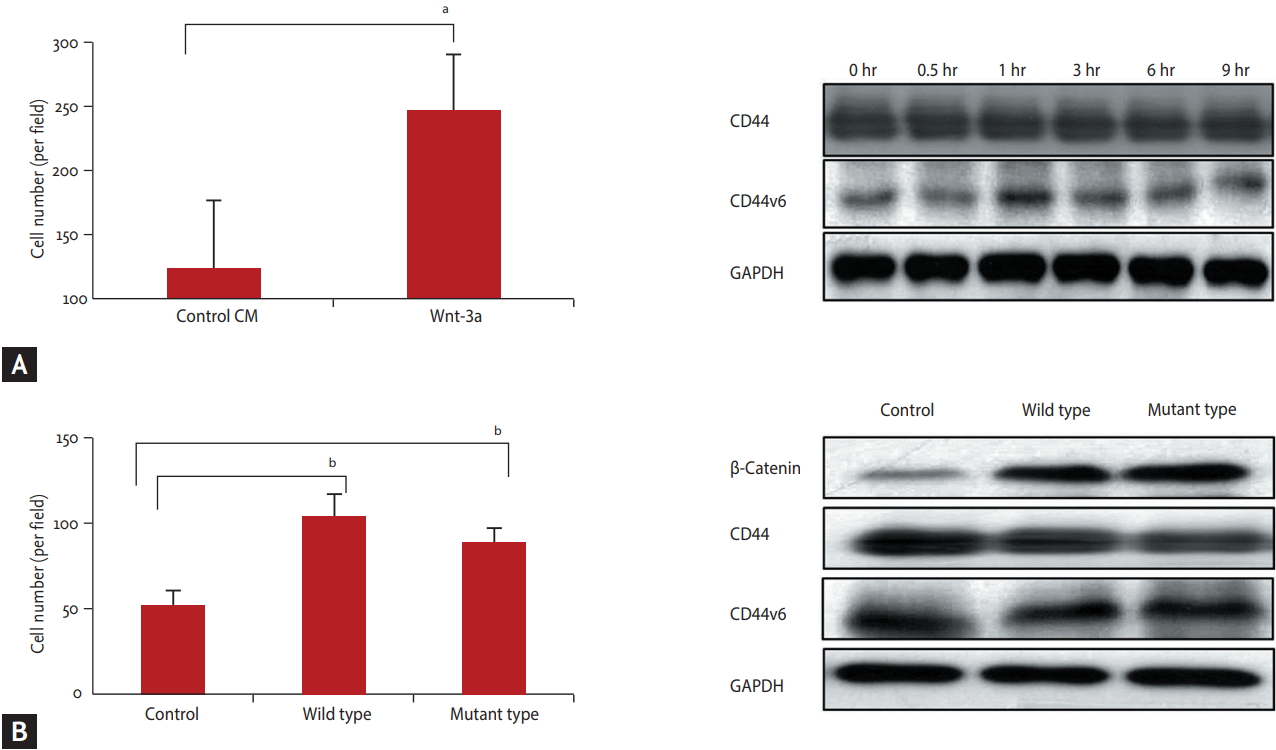
The Panc-1, the low ╬▓-catenin expressing cell line was incubated in Wnt-3a conditioned media and then tested with the cell invasion assay. Invasion was increased after Wnt-3a conditioned media treatment of Panc-1 cell line, low ╬▓-catenin expressing cell line CM (p < 0.05). During treatment with Wnt-3a conditioned media, CD44 or CD44v6 expression was not changed (Fig. 3A). After transfection of ╬▓-catenin into Panc-1 cell line, cell invasion was increased, after transfection of both wildtype and mutant ╬▓-catenin (p = 0.01) (Fig. 3B). It was not associated with CD44 and CD44v6 expression.
Immunohistochemical staining in human tissue
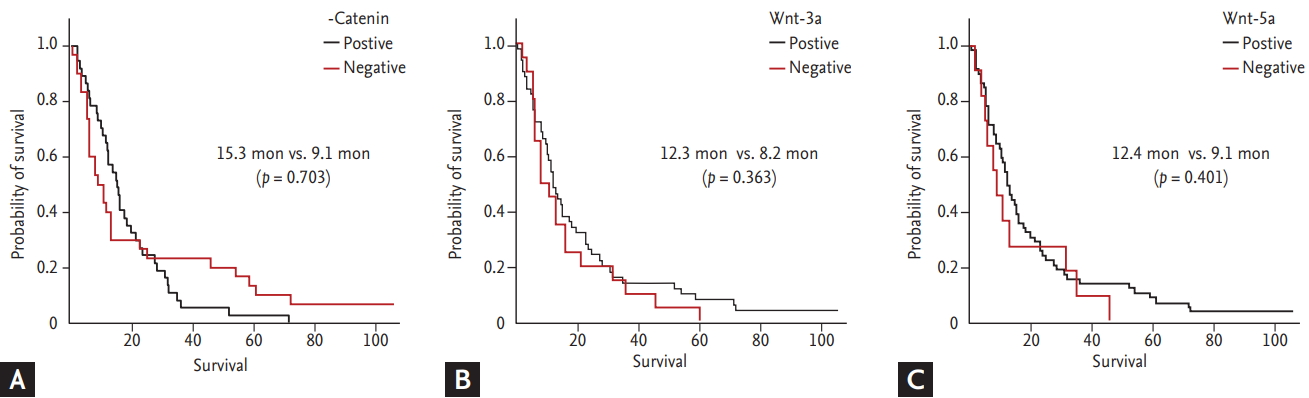
We analyzed the association between Wnt protein expression and the presence of lymph node or development of organ metastasis in the follow-up period. There was no significant correlation with initial stage or lymph node involvement, but Wnt-3a expression was correlated with development of distant organ metastasis after curative surgery (p = 0.029) in the follow-up period (Table 1). However, there was no difference in survival outcome according to Wnt expression (p = 0.703, p = 0.363, and p = 0.401 for ╬▓-catenin, Wnt3-a, and Wnt-5a, respectively) (Fig. 5).
DISCUSSION
Pancreatic cancer is known to be a highly aggressive cancer because of early metastasis, leading to death; however, its mechanism is still not understood. The K-ras mutation is commonly found in 90% of pancreatic cancers and it can activate the Wnt/╬▓-catenin signaling pathway, involved in up-regulation of epithelial mesenchymal transition stimulation [11]. In previous studies, it was reported that Wnt/╬▓-catenin may play a role in cancer invasion or metastasis in hepatocellular cancer and biliary tract cancer [12,13].
In the present study, we tried to determine the association between Wnt/╬▓-catenin and cell invasion. We also investigated the expression of CD44 and CD44 variant, well-known invasion related proteins in pancreatic cancer to identify the relation with Wnt/╬▓-catenin. The ╬▓-catenin expressing cell line, BxPC3 cell line, showed high invasiveness compared to two ╬▓-catenin non-expressing cell lines. These two ╬▓-catenin non-expressing cell lines were not invasive even with high expression of CD44 and CD44 variant. We enhanced or suppressed ╬▓-catenin expression in each cell line to determine if the change in ╬▓-catenin expression affects cell invasion. On adding the Wnt-3a conditioned media to the low ╬▓-catenin expressing cell line; ╬▓-catenin expression was transiently increased, as shown in Fig. 1. When ╬▓-catenin expression was increased, cell invasion of PANC1 was increased, without any change in the CD44 or CD44 variant expression. ╬▓-Catenin was transfected into the PANC1 cell line for producing a transient ╬▓-catenin expressing cell line. We used wild-type and activated ╬▓-catenin to determine if the type of ╬▓-catenin plays a specific role in cell invasion. In our study, the type of ╬▓-catenin did not play a critical role in invasion. It was also not associated with CD44 or CD44 variant expression. Considering that both types of ╬▓-catenin expression significantly increased cell invasiveness, Wnt/╬▓-catenin activation itself can be associated with cell invasion and metastasis, but it may not be associated with CD44 activation.
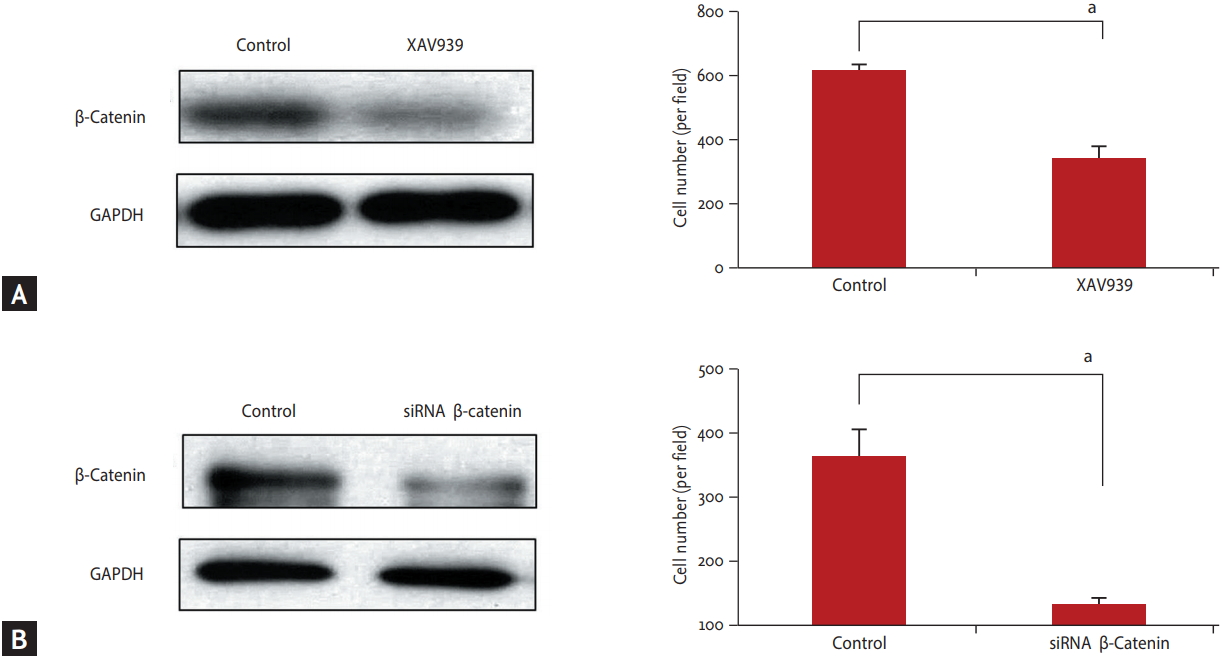
XAV939, a potent tankyrase (TNKS) inhibitor, deregulates the Wnt/╬▓-catenin signaling pathway in various cancers by stabilizing axin, without affecting nuclear factor kappa B or transforming growth factor-╬▓ [14]. We treated BxPC3 cell line with XAV939 to inhibit ╬▓-catenin activity. As shown in Fig. 4, cell invasiveness was significantly decreased during ╬▓-catenin expression. A similar result was found in our experiment with siRNA for ╬▓-catenin. All these data indicated that the Wnt/╬▓-catenin signaling pathway can be involved in cell invasion and metastasis in pancreatic cancer.
In a previous study, the Wnt/╬▓-catenin signal pathway was found to be involved in cancer invasion and metastasis through various mechanisms in solid tumor [5,15]. Pai et al. [7] suggested that the aberrant Wnt signaling pathway contributed to progression of pancreatic cancer by promoting mucin4 (MUC4). In one report, inhibition of Wnt/╬▓-catenin caused regression of tumor growth in a mouse model [16]. However, we investigated the role of ╬▓-catenin by direct enhancement or inhibition of ╬▓-catenin, without using the Wnt signaling pathway related protein unlike the previous trials. This suggested that the target for ╬▓-catenin, not for any step of the Wnt/╬▓-catenin effector pathway can control invasion or metastasis in pancreatic cancer.
We also investigated the association between Wnt protein expression and the development of metastasis in human tissue. In our study, Wnt-3a protein expression showed a correlation with metastasis, but not ╬▓-catenin. First, the Wnt signaling pathway has been known to be activated by two pathways, ╬▓-catenin dependent pathway (canonical pathway) and ╬▓-catenin independent pathway (non-canonical pathway), and the Wnt-3a, in the upper stream, activates both pathways [17]. In the previous study, the non-canonical pathway as well as the canonical pathway was related to cell growth, migration, or metastasis [18-20]. The Wnt-3a may regulate cancer invasion or metastasis through non-canonical pathway, independent of ╬▓-catenin. Second, we analyzed a small number of patients with different stages in the present study; thus, we could not adequately confirm the in vitro data.
There are some limitations to our study; therefore, our results should be interpreted with caution. First, we performed the experiments at a cell line level, and therefore, an in vivo test is required to support our hypothesis. Second, we did not confirm any interaction with other metastasis related protein except CD44. Therefore, we could not explain how Wnt/╬▓-catenin acts on metastasis. The Wnt/╬▓-catenin signaling pathway can interact with many signaling pathways for tumor and tumor microenvironment as a ŌĆ£controller of the signaling pathway.ŌĆØ To define the mechanism for metastasis in pancreatic cancer, we should perform further research considering this point.
In spite of these limitations, our research is worthy as it provides information about the potential role of Wnt/╬▓-catenin associated with metastasis in pancreatic cancer. We also observed the same result in human tissue, and not only in cell line. Based on our result, Wnt/╬▓-catenin can be another potential target to control the progression in pancreatic cancer.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print





