


INTRODUCTION
Graft-versus-host disease (GVHD) is the most severe complication of allogeneic hematopoietic stem cell transplantation (HSCT). GVHD is characterized by an imbalance between the effector and regulatory arms of the immune system, which results in the overproduction of inflammatory cytokines. Several studies have demonstrated that CD4+CD25ŌĆō conventional T cells (Tcons) are critical for the development of GVHD, since depletion of T cells from donor bone marrow (BM) effectively prevents acute GVHD; thus, increasing animal survival rates in murine studies [1,2]. Recently, these promising results have been translated into clinical applications; CD4+CD25+Foxp3+ regulatory T (Treg)-based cellular therapy is considered as a powerful approach for both acute and chronic GVHD prevention.
Treg cells are not a homogenous cell population, but can be divided into at least two subpopulations according to CD62L expression. CD62L (L-selectin) is a member of the selection adhesion molecule family, and is an important T cell homing receptor, as well as a marker for T cell development [3]. The CD62L+ subset of cells is a more potent suppressor than the CD62LŌĆō population or unfractionated CD4+CD25+ Treg cells; in addition, it can be expanded far more easily in culture, and is more responsive to chemokine-driven migration to secondary lymphoid organs [4].
Despite these promising results, there are several factors that limit the clinical applications of GVHD therapy. Several groups are investigating the clinical utility of ex vivo expansion of donor-derived Treg cells, to increase their number, because Treg cells are a rare cell population; others are improving culturing strategies to enhance Treg cell function. Moreover, in terms of actual clinical performance, it is difficult to request another donation of an unrelated donorŌĆÖs blood following HSCT for the purpose of generating Treg cells. Brunstein et al. [5] recently demonstrated the safety and clinical efficacy of administration of third-party cord blood-derived Treg cells after a primary cord blood transplantation. Therefore, third-party-derived Treg cells are particularly suitable for such studies, as they can be prepared in advance and then banked for further use.
Several studies have demonstrated that Treg cells from different sources, such as a donor, recipient, or third-party, have been tested separately in preclinical and clinical transplantation studies, but no comparison among these three types of Treg sources has been systematically reported simultaneously. In the present study, we used a mouse model to test the efficacy of donor, host, or third-party-derived Treg cells.
METHODS
Mice
C57BL/6 (H-2b), BALB/c (H-2d), and DBA1J (H-2q) mice, 8 to 10 weeks old, were purchased from Orient (Seongnam, Korea). Mice were maintained under specific pathogen-free conditions in an animal facility with controlled humidity (55% ┬▒ 5%), light (12/12-hour light/dark), and temperature (22┬░C ┬▒ 1┬░C). The air in the facility was passed through a HEPA filter system designed to exclude bacteria and viruses. Animals were fed mouse chow and tap water ad libitum. The protocols used in this study were approved by the Animal Care and Use Committee of The Catholic University of Korea (2010-0204-02).
Bone marrow transplantation and acute GVHD induction
Recipient mice (BALB/c, H-2d) were irradiated with 800 cGy and injected intravenously (IV) with 5 ├Ś 106 T cell-depleted bone marrow cells (TCD-BM) and 5 ├Ś 106 CD4+CD25ŌĆō splenic T cells from donor mice (C57BL/6, H-2b). Control groups were comprised of irradiated mice receiving only 5 ├Ś 106 TCD-BM cells (which did not induce GVHD). Survival after bone marrow transplantation (BMT) was monitored daily, and the degree of clinical GVHD was assessed weekly using a system that scored changes in five clinical parameters: weight loss, posture, activity, fur texture, and skin integrity.
Treg cell generation
To obtain Treg cells, isolated CD4+ T cells from donors (C57BL/6), recipients (BALB/c) and third parties (DBA1J) were cultured with anti-CD3 (1 ╬╝g/mL), anti-CD28 (1 ╬╝g/mL), human recombinant transforming growth factor (5 ng/mL) and retinoic acid (100 ╬╝M) for 3 days. The expanded induced Treg cells were then sorted by flow cytometry to obtain a ~90% pure CD4+CD25+CD62L+ population [6].
Treg cell therapy
Mice were injected IV with 5 ├Ś 105 Treg cells derived from one of a donor, host or third-party, after BMT (BMT + day 1). Control mice received IV injections of an equal volume of phosphate-buffered saline (PBS) (Gibco, Carlsbad, CA, USA) at the same time points. Donor Treg, host Treg, and third-party Treg refer to donor mice-derived Treg cell, host mice-derived Treg cell, and third party mice derived Treg cell, respectively.
Histopathological analysis of acute GVHD
Survival after BMT was monitored daily, and the degree of clinical GVHD was assessed weekly using a scoring system that sums changes in five clinical parameters: weight loss, posture, activity, fur texture, and skin integrity. Mice were killed at day 15 after BMT for blinded histopathological analysis of GVHD targets (skin, liver, and small and large intestine) [7]. Organs were harvested, cryo-embedded, and subsequently sectioned. Tissue sections were fixed in 10% buffered formalin and stained with hematoxylin and eosin for histological examination.
Flow cytometry
Mononuclear cells were immunostained with various combinations of the following fluorescence-conjugated antibodies: intercellular adhesion molecule 1 (ICAM-1), cytotoxic T lymphocyte antigen-4 (CTLA-4), programmed death-1 (PD-1), inducible costimulator (ICOS), CD103, CD25, CD4, Foxp3, interleukin (IL)-17, interferon-╬│ (IFN-╬│), IL-4, IL-10, and IL-6. These cells were also intracellularly stained with the following antibodies: IL-4, IL-6 (BD Biosciences, Franklin Lakes, NJ, USA), IL-10 (BioLegend), IL-17, and Foxp3 (eBioscience, San Diego, CA, USA). Before intracellular staining, the cells were restimulated for 4 hours with 25 ng/mL PMA and 250 ng/mL ionomycin in the presence of GolgiStop (BD Biosciences). Intracellular staining was conducted using an intracellular staining kit (eBioscience) according to the manufacturerŌĆÖs protocol. Flow cytometric analysis was performed on a FACS LSRFortessa (BD Biosciences).
ELISA assay for cytokine levels
Concentrations of IL-4, IL-21, IL-10, IFN-╬│, IL-6, and IL-17 were measured using a sandwich ELISA as follows. Anti-mouse IL-4, IL-21, IL-10, IFN-╬│, IL-6, or IL-17 monoclonal antibodies (R&D Systems, Minneapolis, MN, USA) were added to a 96-well plate (Nunc, Roskilde, Denmark) and incubated overnight at 4┬░C. Wells were blocked with blocking solution (PBS containing 1% bovine serum albumin and 0.05% Tween 20) for 2 hours at room temperature. Test samples and standard recombinant mouse IL-4, IL-21, IL-10, IFN-╬│, IL-6, and IL-17 (R&D Systems) were added to separate wells and the plate was incubated at room temperature for 2 hours, after which it was washed. Biotinylated IL-4, IL-21, IL-10, IFN-╬│, IL-6, and IL-17 polyclonal antibodies (R&D Systems) were added, and the reaction was allowed to proceed for 2 hours at room temperature. The plate was washed, ExtrAvidin-alkaline phosphatase (1:2,000 dilution; Sigma-Aldrich, St. Louis, MO, USA) was added, and the reaction was allowed to proceed for an additional 2 hours. The plate was washed and 50 ╬╝L p-nitrophenyl phosphate disodium salt (Pierce Chemical Co., Rockford, IL, USA) (diluted to 1 mg/mL in diethanolamine buffer) was applied. Experiments were performed according to the manufacturerŌĆÖs instructions.
RESULTS
Phenotypes of ex vivo expanded Treg cells according to Treg source
Recent studies revealed the importance of diverse patterns of homing receptors for the appropriate tissue distribution and function of Treg cells. Three groups of retinoic acid-induced CD4+CD25+CD62L+ Treg cells, analyzed by flow cytometry, showed > 96% purity and positive surface staining for several phenotypic Treg markers, including CTLA-4 (Fig. 1A), ICOS (Fig. 1B), ICAM-1 (Fig. 1C), CD103 (Fig. 1D), and PD-1 (Fig. 1E). However, DBA1J (third-party)-derived Treg cells showed weakly positive surface staining for ICOS, CTLA-4 and PD-1 as compared with C57BL/6 (donor) [8] and BALB/c (host)-derived Treg cells [9].
Regardless of Treg source, CD4+CD25+CD62L+ Treg cells are protective for lethal acute GVHD
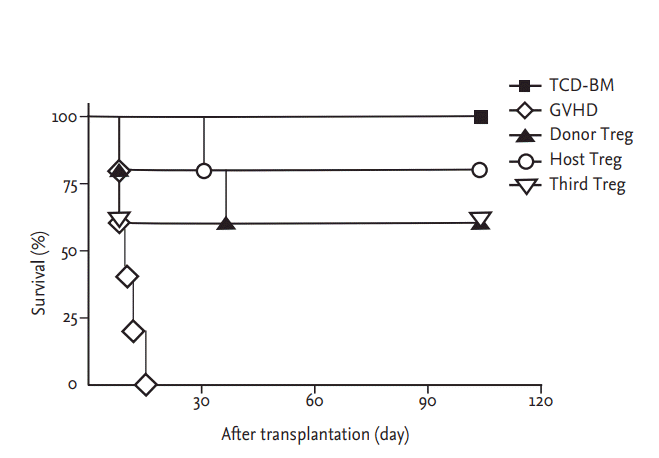
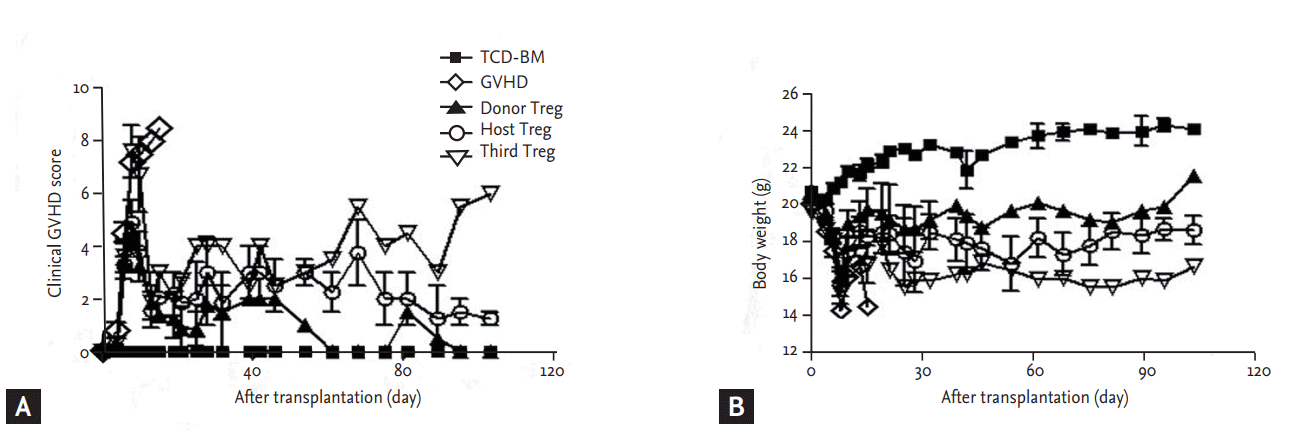
We established a model of acute GVHD following complete major histocompatibility complex (MHC)-mismatched HSCT, to assess the effects of ex vivo expanded Treg cells of various origin on GVHD, engraftment, and immune reconstitution. In this model, 5 ├Ś 106 TCD-BM cells from C57BL/6 (H-2b) mice were injected into lethally irradiated (total body irradiation; 800 cGy) B/c (H-2d) recipient mice to establish hematopoiesis. Co-transfer of 5 ├Ś 105 donor CD4+CD25ŌĆō Tcons resulted in lethal GVHD with a median survival of 10 to 20 days. Infusion of CD4+CD25+CD62L+ Treg cells at a 1:1 ratio to Tcons protected animals from GVHD-related mortality. In these experiments, irradiated BALB/c (H-2d) mice were divided into five groups: mice in group 1 received TCDBM cells from C57BL/6 mice; in group 2, in addition to TCD-BM cells, mice also received donor Tcons to induce GVHD; mice in groups 3 to 5 were transplanted as in group 2 and also received IV infusions with donor-, host-, or third-party-derived CD4+CD25+CD62L+ T cells on day 1 post-transplantation. Recipient mice were monitored for clinical scores of GVHD, weight, and survival. As expected, none of the animals from group 1 died after transplantation, whereas all the animals from group 2 died within 20 days; no significant differences in survival rates were observed among groups 3 to 5 (Fig. 2) and GVHD clinical scores indicated that donor-derived Treg cell-treated mice did not develop clinical signs of GVHD for > 100 days as compared with host-derived Treg cell-treated and third-party-derived Treg cell-treated mice (Fig. 3A). Also, mice receiving an infusion of donor-derived Treg cells gained a significant amount of weight (Fig. 3B). Notably, the improved GVHD scores of mice in group 3 were consistent with a continued health status until at least day 100 after HSCT.
Regardless of Treg source, CD4+CD25+CD62L+ Treg cell therapy reduces the histopathological manifestations of acute GVHD
Histological analysis of skin from the group of mice with acute GVHD showed scattered eosinophilic degradation (apoptosis) and slight vacuolation of individual epidermal cells. These corresponded to of a score of 8 according to the criteria used [10]. The other Treg cell-treated groups (donor, host, and third-party Treg cells) showed reduced histologic scores, but there were no significant differences among them (Fig. 4A). Histological grading of GVHD of the small intestine showed effacement and blunting of the villous architecture, mucous cell depletion and sloughing of epithelial cells with patchy mucosal ulceration, yielding an acute GVHD histologic score of 2. These outcomes were improved with the addition of donor Treg cells in the group of mice with GVHD. Histological analysis of the liver showed a slight infiltration of lymphoid cells in the portal space and remarkable changes in the bile duct epithelium. Therefore, the total histological scores of each host, third-party, and donor Treg group were lower than that of GVHD group, yet the total score of donor Treg was the lowest. (Fig 4B).
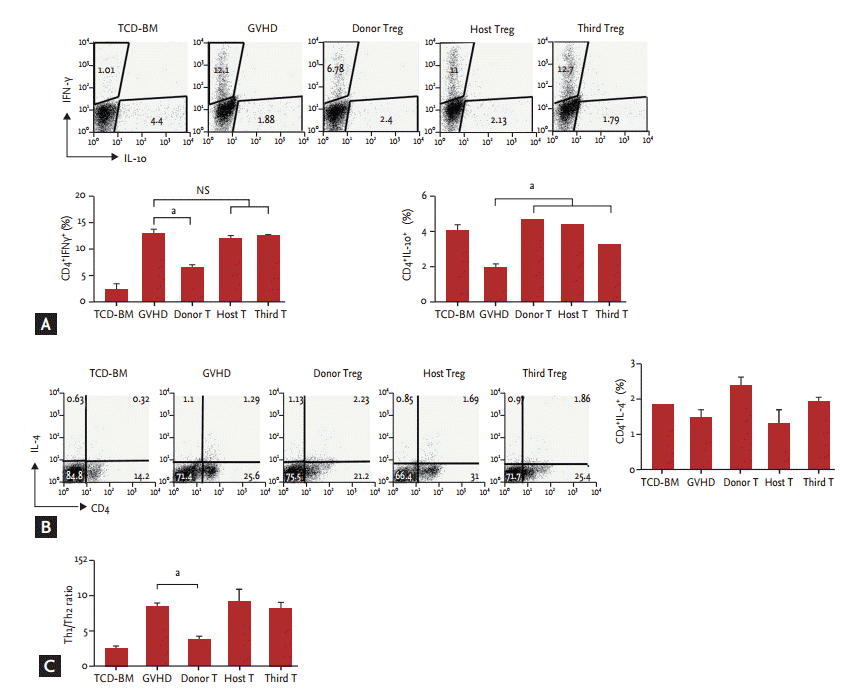
Third-party-derived CD4+CD25+CD62L+ Treg cells did not modulate the balance of Th1 and Th2 cells in lethal acute GVHD induced by CD4+CD25ŌĆō T cells
To confirm the immunoregulatory function of donor Treg, host Treg, and third-party Treg cell therapy groups relative to T-helper 1 (Th1) and Th2 cells, flow cytometry was used to measure cytokine expression. Mice of the host, third-party Treg cell and GVHD control groups produced similar or slightly higher levels of IFN-╬│, whereas those of donor Treg cell-treated GVHD mice showed markedly decreased production of inflammatory cytokines. However, three groups of Treg cell-treated GVHD mice displayed increased IL-10 expression in the spleen compared with GVHD control mice (Fig. 5A). Differences in IL-4 levels between the groups was not statistically significant (Fig. 5B). Analyses performed on the spleens of donor Treg cell-treated GVHD mice showed that the Th1/Th2 ratio was the lowest among all of the groups (Fig. 5C). Thus, third-party-derived Treg cell therapy did not affect Th1/Th2 cytokine production. However, donor-derived Treg cell therapy markedly alters the balance between Th1 and Th2 cells in GVHD mice, resulting in an altered disease course.
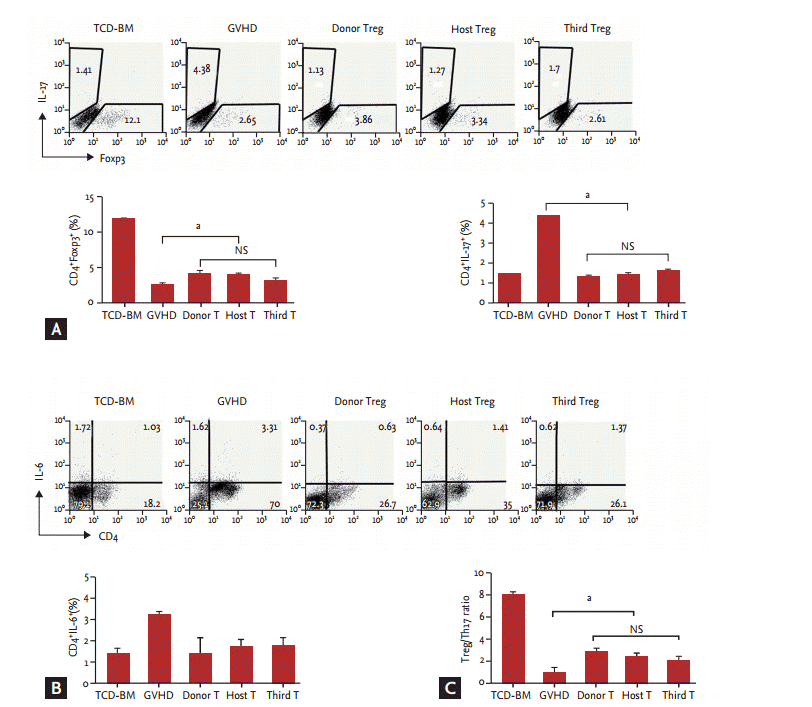
Third-party-derived CD4+CD25+CD62L+ Treg cell therapy provides an immunomodulatory effect associated with reciprocal regulation of 17+ Th17/Treg cells
Foxp3 expression in the three groups of CD4+CD25+CD62L+ Treg cell therapy was significantly higher than in the control GVHD group, whereas expression of IL-17 and IL-6 in the three CD4+CD25+CD62L+ Treg cell therapy groups was significantly lower than in the control group. However, among the three Treg cell therapy groups, no difference was observed in the expression of Foxp3, IL-17, or IL-6 (Fig. 6A and 6B). Th1 and Th17 cells in murine GVHD were associated with evidence of severe GVHD, and in situ quantification of the Treg/Th17 ratio was a specific marker for GVHD. The Treg/Th17 cell ratio was significantly higher in spleens from mice treated with the three types of CD4+CD25+CD62L+ Treg cells, as compared with the control GVHD group, in the mouse model of GVHD (Fig. 6C). Taken together, these data might explain why third-party-derived Treg cell therapy equally inhibits acute GVHD in pathogenic CD4+CD25ŌĆō donor T cell-treated mice compared with donor- or host-derived Treg cell-treated groups.
DISCUSSION
Preclinical murine transplantation models have convincingly established that Treg cells have the capacity to prevent alloreactive T-cell responses in experimental models of both acute and chronic GVHD [11,12]. For example, hematopoietic stem cell grafts with a higher content of Treg cells have been correlated with less acute GVHD [13]. Likewise, more rapid Treg reconstitution is associated with less acute GVHD, whereas patients with delayed Treg cell recovery have a higher likelihood of developing GVHD [14].
Adoptive transfer of in vitro-differentiated induced Treg cells, along with BM grafts containing alloreactive donor T cells, did not result in any significant protection against lethal acute GVHD [15,16], although one study did demonstrate its efficacy in a lupus-like chronic GVHD model [17]. A major reason for the lack of observed protection in the acute GVHD models was the limited in vivo survival of these cells, accompanied by instability of Foxp3 expression that resulted in an early loss of suppressive function post-transplantation [15,18]. However, increased numbers of CD62L+ Treg cells in grafts have been found to correlate with reduced GVHD incidence [19], which is likely due to the ability of these cells to enter secondary lymphoid tissue, where allorecognition by donor T cells and acute GVHD initiation occur. This is consistent with reported data from murine models of acute GVHD, in which the CD62L+ Treg cell population suppresses acute GVHD to a greater extent than the corresponding CD62Llow population [20,21]. Somewhat lacking, but we showed that ex vivo-expanded isolated third-party-derived CD4+CD25+CD62L+ Treg cells showed similarly potent suppressive activity to that of donor- or host-derived cells.
The present study demonstrates that ex vivo expanded third-party-derived CD4+CD25+CD62L+ Treg cells are a valuable and useful alternative to Treg cells derived from the same donor or host as the Tcon cells, to effectively suppress acute GVHD. Our results showed that the major limitation to the adoptive transfer of third-party-derived Treg cells was that this therapy resulted in slightly worse clinical symptoms in vivo in comparison with donor-derived Treg cell therapy, which were present for immunomodulatory functions during alloreactive T cells. However, both third-party-derived CD4+CD25+CD62L+ Treg cell therapy and donor-derived CD4+CD25+CD62L+ Treg cell therapy provide an immunomodulatory effect associated with reciprocal regulation of Th17 and Treg cells after allogeneic BMT.
Recently, clinical studies have been published showing that generation of third-party-derived cord blood Treg cells via the ex vivo expansion strategy has the potential to provide a readily available, clinically relevant, ŌĆ£off-the-shelfŌĆØ cellular therapy to prevent GVHD and improve transplant outcomes [22]. Although the clinical trials using Treg cells to prevent acute GVHD revealed no obvious safety issues, there are still several concerns regarding this application in the clinic. One disadvantage of administration of third-party cord blood-derived Treg cells is possible relapse and increased incidence of infectious complications caused by the suppressor function of Treg cells. However, in previous clinical trial studies, no such complications were observed [22]. This report demonstrates that adoptive transfer of third-party-derived CD4+CD25+CD62L+ Treg cells is a promising alternative to donor-derived Treg-based cellular therapy, which could result in reduction of GVHD incidence and improved transplant outcomes.


 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print





