


INTRODUCTON
The metabolic complications of obesity include increased risks of type 2 diabetes mellitus (T2DM), cardiovascular diseases (CVDs), sleep apnea, cancer, fatty liver, and other diseases. It is evident that obesity promotes a state of chronic inflammation in white adipose tissue (WAT), which, in turn, leads to insulin resistance. Adipose tissues secrete adipokines that play major roles in metabolism, immunity, and inflammation. Resistin is primarily secreted by adipocytes in rodents and was initially proposed as a link between obesity and insulin resistance [1]. In humans, however, resistin is mainly secreted by macrophages [2], which suggests that resistin is linked to inflammation. Recently, obesity, T2DM, and CVD have been recognized as chronic inflammatory disorders that may be connected to proinflammatory cytokines and various adipokines, including resistin. Thus, the present review will discuss the biology of resistin in rodents and humans with a focus on the evolving roles that resistin plays in inflammation and obesity-related diseases such as atherosclerosis, CVD, atherogenic dyslipidemia, and hypertension in humans.
RESISTIN IN RODENTS AND HUMANS
Resistin was initially identified in murine white adipocytes and is characterized by the presence of 10 to 11 cysteine residues [3]. The expression patterns of resistin in rodents and humans differ considerably. While murine resistin is expressed primarily by adipocytes, human resistin is predominantly expressed by peripheral blood mononuclear cells (PBMC) and macrophages [2]. The lack of resistin expression in human adipocytes may be due to the absence of a genomic binding site for nuclear receptor peroxisome proliferator-activated receptor γ, which normally controls the expression of the resistin gene (retn) in mouse adipocytes [4]. Human resistin shares only 59% of the sequence homology of murine resistin at the amino acid level and the resistin genes in rodents and humans have markedly divergent promoter regions, which suggests that there are differences in regulation and function. Resistin expression in mouse adipocytes is stimulated by high glucose levels but suppressed by tumor necrosis factor α (TNF-α) while human resistin is robustly induced by various inflammatory stimuli, including lipopolysaccharide (LPS), TNF-α, interleukin 6 (IL-6), IL-1β, and resistin itself, in PBMC and macrophages [5].
There are heightened circulating resistin levels in diet-induced obese and ob/ob mice but decreased levels following rosiglitazone treatment [1]. As in rodents, thiazolidinedione treatment downregulates the expression of human resistin in macrophages and reduces serum resistin levels. In mice, circulating resistin exists in a disulfide-linked hexamer or a smaller trimer that exhibits greater bioactivity under conditions of lowered hepatic insulin sensitivity. In humans, resistin circulates in trimeric and oligomeric forms of which the oligomeric form appears to have a more potent effect on the stimulation of proinflammatory cytokines [6]. Interestingly, the bioactivity of resistin is augmented by a physical interaction between resistin and heparanase [7].
ROLE OF MURINE RESISTIN IN INSULIN RESISTANCE
Resistin is a mediator of insulin resistance in rodents. For example, circulating resistin levels are elevated in obese mice and the administration of recombinant resistin impairs insulin action in normal mice. The central and peripheral administration of recombinant resistin and the transgenic overexpression of resistin induce hepatic insulin resistance in mice and, similarly, a variety of cells and tissues in rodents are responsive to resistin. The treatment of 3T3-L1 adipocytes, murine cardiomyocytes, and cultured skeletal muscle cells with murine resistin decreases insulin-stimulated glucose uptake into the cells [8] and resistin induces hepatic insulin resistance in primary rat hepatocytes and a mouse model with a high liver-specific expression of resistin [9]. Conversely, the neutralization of resistin with anti-resistin antibodies improves insulin sensitivity in diet-induced obese mice and the knockdown or deletion of resistin increases hepatic insulin sensitivity in mice on a high-fat diet [5].
The precise mechanisms by which murine resistin exerts its biological effects are not yet fully understood because the resistin receptor has yet to be characterized. However, recent studies have provided several potential candidates for the murine resistin receptor, including an isoform of decorin known as tyrosine kinase-like orphan receptor-1 (ROR1). Similarly, it has been proposed that a cleavage product of decorin, which is known as δ-decorin and lacks a glycanation site, is a functional receptor of murine resistin in adipose progenitor cells [10]. In fact, δ-decorin, as a functional resistin receptor, may regulate WAT expansion by modulating cell proliferation and migration as well as adipocyte differentiation in mice. Additionally, it has been shown that the interaction of mouse resistin with ROR1 results in the inhibition of ROR1 phosphorylation, which modulates the phosphorylation of extracellular signal-regulated kinases (ERKs) 1/2 and regulates the expression of suppressor of cytokine signaling 3 (SOCS3) and glucose transporter 4 in 3T3-L1 preadipocytes [11].
Murine resistin also decreases the phosphorylation of 5’ adenosine monophosphate-activated protein kinase (AMPK) in the liver, skeletal muscle, and WAT [12,13] while the inhibition of resistin leads to an increase in the phosphorylation of AMPK [14]. Furthermore, resistin interferes with multiple steps of the insulin signaling cascade, including the phosphorylation of insulin receptor substrates and the activations of phosphatidylinositol-3-kinase and protein kinase B/Akt in the rodent liver, muscle, and WAT [13-15]. Resistin treatment in rodents also induces the expression of SOCS3, which is a known inhibitor of insulin signaling, in the liver, muscle, and WAT [15]. Despite differences in the mechanisms of action underlying the effects of resistin in rodents and humans, the evidence indicates that murine resistin is linked to insulin resistance and has led to advances in research aimed at understanding the biology of human resistin.
ROLE OF HUMAN RESISTIN IN INFLAMMATION
Human resistin plays a major regulatory role in the inflammatory response [16] during which macrophages, PBMC, and vascular cells are the primary targets of resistin [17]. Resistin upregulates the expression of proinflammatory cytokines such as TNF-α, IL-6, IL-12, and monocyte chemoattractant protein (MCP)-1 in PBMCs, macrophages, and hepatic stellate cells via the nuclear factor-κB (NF-κB) pathway [18]. Resistin expression has also been identified in the non-adipocyte stromal vascular fraction of WAT, fibrotic livers, and atherosclerotic lesions [19]. Moreover, circulating resistin levels are correlated with inflammatory and fibrinolytic markers such as C-reactive protein (CRP), TNF-α, and IL-6 in in the general population and in individuals with T2DM, coronary atherosclerosis, chronic kidney disease, rheumatoid arthritis, and/or sepsis [6]. Plasma resistin levels are also related to disease severity in patients with severe sepsis or acute pancreatitis and are predictive of worse outcomes in non-septic but critically ill patients [5].
ROLE OF HUMAN RESISTIN IN ATHEROSCLEROSIS
In endothelial cells, human resistin increases the expressions of various proinflammatory factors, including MCP-1, endothelin-1, and matrix metalloproteinases, as well as adhesion molecules such as intercellular adhesion molecule-1, vascular cell adhesion molecule-1, and P-selectin [20,21]. Human resistin also increases the proliferation and migration of human endothelial cells and vascular smooth muscle cells (VSMC) and increases endothelial permeability, which promotes endothelial cell/monocyte adhesion and infiltration [22]; these actions are mediated by the ERK and p38 mitogen-activated protein kinase signaling pathways [20]. Furthermore, resistin inhibits endothelial nitric oxide synthase via oxidative stress in human endothelial cells [23], promotes foam cell formation in human macrophages, induces a prothrombotic phenotype in human endothelial cells [17], and induces platelet activation by increasing P-selectin expression [24]. Taken together, these findings suggest that human resistin might play an important regulatory role in the modulation of interactions between endothelial cells, monocytes/macrophages, and VSMC in the pathogenesis and progression of atherosclerosis [25].
ROLE OF HUMAN RESISTIN IN INSULIN RESISTANCE
Given the association between murine resistin and insulin resistance, numerous clinical studies have examined the relationship between circulating resistin levels and insulin resistance in obese people with and without diabetes [6]. While some studies have shown positive correlations between resistin and insulin resistance, other studies have failed to identify significant changes in resistin levels in patients with insulin resistance and/or T2DM [8,25]. However, these discrepancies may be attributed to small sample sizes, population differences, disease status, and the use of non-standardized assays for measuring resistin [26].
To investigate the hypothesis that human resistin contributes to insulin resistance, Qatanani et al. [27] produced a ‘humanized resistin’ transgenic mouse model in which human resistin is primarily expressed in PBMC and macrophages but murine resistin is lacking. When these transgenic mice were fed a high-fat diet, they developed WAT inflammation and insulin resistance in the muscle and adipose tissues, which suggests that human resistin may be a link between the inflammatory environment and insulin resistance [27]. However, in this transgenic mouse model, the expression of human resistin in macrophages was constitutive and not induced by inflammatory stimuli. Therefore, it was unclear whether resistin induced by inflammatory signals could lead to insulin resistance in humans. To address this question, the Lazar laboratory generated a bacterial artificial chromosome transgenic mouse that lacked murine resistin but expressed human resistin, which was referred to as the Hum-Retn mouse [28]. Hum-Retn mice exhibit serum resistin levels similar to the normal range for humans. Additionally, in contrast to the earlier macrophage-driven resistin transgenic mice, human resistin levels in Hum-Retn mice show a marked increase in response to LPS treatment, which is also similar to humans [28]. Moreover, under conditions of chronic endotoxemia, Hum-Retn mice develop hepatic insulin resistance that is accompanied by inflammation in the liver and skeletal muscle, which supports the idea that resistin is involved in the pathophysiology of inflammation-induced insulin resistance in humans [28].
In line with this observation, it has been shown that human resistin inhibits insulin-stimulated glucose uptake in isolated mouse cardiomyocytes [29]. Resistin also blunts insulin signaling in myocytes via the suppression of myogenesis and stimulation of the proliferation of myoblasts in C1C12 myotubes [30], and induces insulin resistance via AMPK-dependent and AMPK-independent mechanisms in HepG2 hepatocytes [31]. Recently, prospective case-control studies have found that individuals with elevated baseline levels of resistin have a significantly higher risk of developing T2DM, even after adjusting for other risk factors [5,32].
POTENTIAL CANDIDATES FOR THE HUMAN RESISTIN RECEPTOR
The human resistin receptor remains unknown but Toll-like receptor 4 (TLR4) and adenylyl cyclase-associated protein 1 (CAP1) have recently been proposed as a putative candidates [33,34]. TLR4 serves as a receptor for the proinflammatory effects of recombinant human resistin in human myeloid and epithelial cells [33]. Additionally, recent findings in rats have shown that the direct binding of human resistin to TLR4 in the hypothalamus results in the activation of proinflammatory pathways and insulin resistance, which indicates that TLR4 is a binding site for resistin [35]. CAP1 has also been proposed as the in vivo functional receptor for resistin in human monocytes [34]. Resistin binds directly to CAP1 in monocytes and upregulates cyclic adenosine monophosphate concentrations, protein kinase A activity, and the NF-κB-related transcription of inflammatory cytokines in human monocytes [34]. However, further investigations will be necessary to clarify the relevance of these identified receptors in light of the biological effects of resistin in a variety of metabolic processes.
CLINICAL IMPLICATIONS OF HUMAN RESISTIN IN CVD
An increasing amount of evidence indicates that resistin is associated with coronary and peripheral artery diseases, ischemic stroke, and congestive heart failure [36]. Elevated resistin levels are predictive of coronary atherosclerosis independently of CRP levels in asymptomatic individuals and resistin is a predictor of re-stenosis after coronary artery stenting and major adverse cardiovascular events in patients with coronary artery disease (CAD) [6]. Additionally, elevated resistin levels are an independent predictor of all-cause mortality in Korean patients with acute myocardial infarction [37]. A prospective longitudinal study found that resistin was the only predictor of reduced amputation-free survival in patients with diabetes undergoing lower extremity bypass surgery [36] while a recent analysis investigating T2DM patients of European ancestry from two cross-sectional and two prospective studies revealed that resistin predicts incident major cardiovascular events and all-cause mortality [38]. More recently, it was shown that resistin levels are associated with recurrent coronary events in patients after hospitalization for acute coronary syndrome [39] and an association between resistin and ischemic stroke has been identified in some studies [6]. Similarly, a cross-sectional study from Japan and two nested case-control studies from Europe and the United States demonstrated that resistin is significantly associated with ischemic stroke among healthy individuals [6]. Higher resistin levels are also associated with a poorer prognosis in patients with CAD or atherothrombotic stroke [6]. Taken together, the abovementioned clinical evidence suggests a role for resistin in atherosclerotic disease in humans.
Recent clinical studies have demonstrated that elevated serum resistin levels are associated with the development and severity of heart failure [5]. Likewise, large cohort studies have shown that high resistin is independently associated with new-onset heart failure and that resistin is predictive of adverse cardiac outcomes in patients with heart failure [6]. However, whether human resistin acts as an etiological factor in the development of heart failure remains uncertain. Murine resistin is upregulated in rat cardiomyocytes following mechanical stretch [40] and the overexpression of murine resistin decreases left ventricular contractility and promotes cardiac hypertrophy in rats [41]. Interestingly, pre-treatment with murine resistin protects the heart against ischemia/reperfusion injury in a mouse heart perfusion model [42]. In contrast, pre-conditioning with recombinant human resistin prior to ischemia impairs contractile recovery during reperfusion and stimulates cardiac TNF-α secretion in isolated rat hearts [43]. The role of human resistin in the development of cardiotoxicity has also been explored using Hum-Retn mice [44]. Doxorubicin-induced cardiotoxicity is enhanced in Hum-Retn mice with a 4-fold elevation in serum resistin levels, as compared to control mice that lack human resistin [44]. Moreover, a clinical study found that elevated resistin levels are associated with a decline in ejection fraction in women treated for breast cancer with anthracycline-containing chemotherapy [44].
GENETIC EVIDENCE FOR THE ROLES OF HUMAN RESISTIN IN INSULIN RESISTANCE AND ATHEROSCLEROSIS
Several single-nucleotide polymorphisms (SNPs) in the human resistin gene (RETN) have been associated with elevated resistin levels and up to two-thirds of the variation in serum resistin levels may be attributable to genetic factors [45]. However, the identified variants of the RETN gene are inconsistently related to insulin resistance and atherosclerosis. In a Japanese population, the –638 G>A, –420 C>G, and –358 G>A polymorphisms in the promoter region of the RETN gene are associated with resistin levels while the G/G genotype at SNP –420 in the RETN gene is associated with susceptibility to T2DM and has been correlated with the expression of resistin in monocytes [5,6]. An analysis of 2,078 Japanese subjects revealed that plasma resistin is associated with SNP –420 and is correlated with insulin resistance [46]. In another study of a Korean population, the –420G and –537A alleles were associated with increased resistin levels but not T2DM [47]. In a study investigating Chinese subjects, the presence of the –420G and +62A alleles predicted worse glucose tolerance at the 5-year follow-up assessment [48].
Although the –420G allele that is associated with high resistin levels appears to be related to the risk of diabetes in Asians, data from non-Asian populations are conflicting [5]. An Italian study showed that the presence of the –420 C/G SNP in the RETN gene is associated with increased obesity and metabolic syndrome while an analysis of the Framingham Offspring cohort found that the –420 C>G SNP is not associated with obesity or diabetes but, rather, that SNPs in the 3’ region of RETN are associated with resistin levels [5]. On the other hand, a study of obese Egyptian subjects found that both the –420 C>G and +299 G>A SNPs are significantly associated with resistin levels, obesity, and insulin resistance [49]. In a Chinese population, the –420 C/G SNP in the RETN gene was associated with an increased risk of CAD while some studies of European and Caucasian populations did not find an association between the –420 C/G SNP in the RETN gene and coronary atherosclerosis [6].
ROLE OF HUMAN RESISTIN IN ATHEROGENIC DYSLIPIDEMIA
A large population-based study found a strong positive correlation between circulating resistin levels and fasting serum triglyceride concentrations [50]. In Indian men, serum resistin levels are positively correlated with very low density lipoprotein (VLDL) cholesterol and triglyceride levels but negatively correlated with high density lipoprotein cholesterol levels [51]. In contrast, other studies have shown that resistin is inversely correlated with low density lipoprotein (LDL) cholesterol levels [52,53]. It has been suggested that resistin is positively correlated with atherogenic smaller and denser LDL subclasses in subjects with T2DM [54] and a recent study showed that the treatment of human hepatocytes with resistin potently stimulated the secretion of VLDL apolipoprotein B (apoB) and lipids as well as increased hepatocyte lipid content [55]. Conversely, the removal of serum resistin via immunoprecipitation reduces hepatocyte apoB production [55]. The mechanisms by which resistin increases human hepatocyte VLDL production include the stimulation of apoB100 synthesis and microsomal triglyceride transfer protein activity, the induction of de novo lipogenesis via the SREBP1 and SREBP2 pathways, and the enhancement of apoB100 stability [56].
Resistin increases proprotein convertase subtilisin/kexin type 9 (PCSK9) in human hepatocytes, which, in turn, downregulates the LDL receptor [57] by inducing its degradation [58]. Resistin produces a significant reduction in hepatocyte LDL receptor levels, which mediates its VLDL effects, by inducing cellular PCSK9 expression [55]. Resistin and PCSK9 share a structural homology in the cysteine-rich portion within the C-terminal domain and, thus, it is possible that resistin and PCSK9 form aggregation complexes to target the LDL receptor [59]. In a study investigating a Chinese cohort, plasma resistin levels were significantly related to PCSK9 levels in CAD patients with normal body weight [60]. Taken together, these results indicate that resistin stimulates hepatic VLDL production, increases hepatic lipogenesis, and suppresses LDL receptor levels via increases in hepatocyte PCSK9. Thus, resistin may be a potential target for the treatment of atherogenic dyslipidemia in obese individuals with insulin resistance.
ROLE OF HUMAN RESISTIN IN HYPERTENSION
The central administration of resistin increases renal and lumbar sympathetic nerve activity (SNA) but decreases SNA in brown adipose tissue in rats, which suggests that resistin may contribute to the increased muscle SNA and reduced energy expenditure that is observed in subjects with obesity and diabetes [61]. Although blood pressure in rats is not significantly affected by centrally administered resistin [61], some clinical studies have demonstrated that circulating resistin levels are associated with hypertension in humans [62]. A positive correlation between plasma resistin levels and blood pressure has been observed in subjects both with and without T2DM [63,64]. Plasma resistin levels are increased in the young healthy offspring of individuals with essential hypertension compared to the young healthy offspring of normotensives [65]. Moreover, it has been demonstrated that higher plasma resistin levels are associated with an increased risk for incident hypertension among women without diabetes or hypertension, which is indicative of the effects of resistin on blood pressure [66]. However, to date, the precise mechanisms by which resistin affects blood pressure remain unclear.
CONCLUSIONS
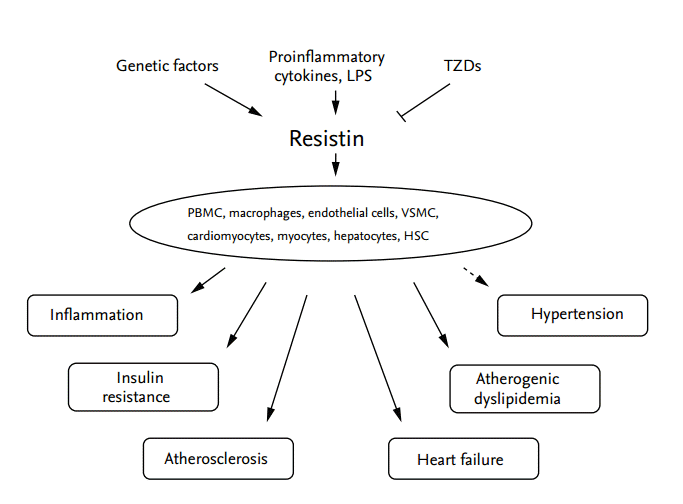
Resistin appears to be a link between obesity and insulin resistance in rodents, and studies of ‘humanized resistin’ transgenic mice have implicated human resistin in the development of inflammation and insulin resistance. Additionally, a growing body of evidence from epidemiological, genetic, and clinical studies indicates that human resistin is involved in the pathogeneses of inflammation, insulin resistance, atherosclerosis, hypertension, and atherogenic dyslipidemia (Fig. 1). Further work is necessary to identify the specific receptor for human resistin, which would lead to delineation of specific signaling pathways and aid in the clarification of the role of resistin in the pathogeneses and treatments of T2DM, dyslipidemia, and CVD.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print





